COMMUNITY &ECOSYSTEM ECOLOGY Light Brown Apple Moth in California: A Diversity of Host Plants and Indigenous Parasitoids XIN-GENG WANG, KARMIT LEVY, NICHOLAS J. MILLS, AND KENT M. DAANE 1 Department of Environmental Science, Policy and Management, University of California, Berkeley, CA 94720 Environ. Entomol. 41(1): 81Ð90 (2012); DOI: http://dx.doi.org/10.1603/EN11160 ABSTRACT The light brown apple moth, Epiphyas postvittana (Walker), an Australia native tor- tricid, was found in California in 2006. A Þeld survey of host plants used by E. postvittana was conducted in an urban region of the San Francisco Bay Area. An inspection of 152 plant species (66 families), within a 23-ha residential community, found E. postvittana on 75 species (36 families). Most (69 species) host plants were not Australian natives, but had a wide geographic origin; 34 species were new host records for E. postvittana. Heavily infested species were the ornamental shrubs Myrtus communis L., Pittosporum tobira (Thunb.) W.T. Aiton, Euonymus japonicus Thunb., and Sollya het- erophylla Lindl. To survey for parasitoids, four urban locations were sampled, with E. postvittana collected from Þve commonly infested plants [M. communis, P. tobira, E. japonicus, Rosmarinus officinalis L., and Genista monspessulana (L.) L.A.S. Johnson]. Twelve primary parasitoid species and two hyperparasitoids were reared; the most common were the egg parasitoid Trichogramma fasciatum (Perkins), the larval parasitoids Meteorus ictericus Nees, and Enytus eureka (Ashmead), and the pupal parasitoid Pediobius ni Peck. Meteorus ictericus accounted for 80% of the larval parasitoids, and was recovered from larvae collected on 39 plant species. Across all samples, mean parasitism was 84.4% for eggs, 43.6% for larvae, and 57.5% for pupae. The results are discussed with respect to the potential for resident parasitoid species to suppress E. postvittana populations. KEY WORDS Lepidoptera, Totricidae, invasive species, host plant use, biological control The light brown apple moth, Epiphyas postvittana (Walker), is a tortricid leafroller that is native to southeastern Australia (Danthanarayana 1975). It has invaded and established in western Australia, New Zealand, Caledonia, HI, the United Kingdom (Suck- ling and Brockerhoff 2010), and, most recently, Cal- ifornia (Brown et al. 2010). The moth is thought to have Australian herbaceous plants as its ancestral hosts (Danthanarayana et al. 1995), but is known to feed on numerous herbaceous plants, ornamental shrubs and trees, and fruit and vegetable crops (Dan- thanarayana 1975, Geier and Briese 1980, Wearing et al. 1991). The long list of recorded host plants includes over 120 species found in Australia, including 23 Aus- tralian natives (Danthanarayana 1975, Geier and Bri- ese 1980), and 250 plant species found in New Zealand (Thomas and Shaw 1982). Epiphyas postvittana may attack over 500 plant species in 363 genera (Suckling and Brockerhoff 2010), and it has become a pest on grapes, apples, caneberries, stone fruits, citrus, and numerous ornamentals in Australia and New Zealand, and is considered to be an important invasive pest for North America and elsewhere (Varela et al. 2008, Brown et al. 2010, Suckling and Brockerhoff 2010). The arrival of E. postvittana in California Þrst was documented in 2006 when the moth was found in Berkeley, CA (Alameda Co.) (Brown et al. 2010). The pest since has been found in 18 Californian counties, with the largest numbers of moths captured in the Central Coast region (Monterey and Santa Cruz coun- ties) and Bay Area (which includes San Francisco, Alameda, and Contra Costa counties) (CDFA 2011). Currently, it is most often found on ornamental plant species in urban areas, with small incursions into some agricultural crops in the Central Coast (CDFA 2011). Commonly infested plants include species from di- verse geographic origins, such as a native Californian manzanita, Arctostaphylos densiflora M.S. Baker; the Mexican orange, Choisya dumosa (Torr.) A. Gray; a Mediterranean native myrtle, Myrtus communis L.; Japanese cheesewood, Pittosporum tobira (Thunb.) W.T. Aiton; and Australian tea tree, Leptospermum laevigatum (Sol. ex Gaertn.) F. Muell. (Mills et al. 2009, Buergi et al. 2011). Although the availability of susceptible host plant species may not limit this pestÕs movement in California, climate may prove to be the greatest delimiting factor. Epiphyas postvittana favors regions with mild summers (Thomson et al. 2010) and in California the pest has not yet been found in the warmer interior valleys, where the hot summers may limit its distribution (Gutierrez et al. 2010) into these 1 Corresponding author, e-mail: [email protected]. 0046-225X/12/0081Ð0090$04.00/0 2012 Entomological Society of America
Transcript
COMMUNITY & ECOSYSTEM ECOLOGY
Light Brown Apple Moth in California: A Diversity of Host Plants andIndigenous Parasitoids
XIN-GENG WANG, KARMIT LEVY, NICHOLAS J. MILLS, AND KENT M. DAANE1
Department of Environmental Science, Policy and Management, University of California, Berkeley, CA 94720
ABSTRACT The light brown apple moth, Epiphyas postvittana (Walker), an Australia native tor-tricid, was found in California in 2006. A Þeld survey of host plants used byE.postvittanawas conductedin an urban region of the San Francisco Bay Area. An inspection of 152 plant species (66 families),within a 23-ha residential community, found E. postvittana on 75 species (36 families). Most (69species) host plants were not Australian natives, but had a wide geographic origin; 34 species werenew host records for E. postvittana. Heavily infested species were the ornamental shrubs Myrtuscommunis L., Pittosporum tobira (Thunb.) W.T. Aiton, Euonymus japonicus Thunb., and Sollya het-erophylla Lindl. To survey for parasitoids, four urban locations were sampled, with E. postvittanacollected from Þve commonly infested plants [M. communis, P. tobira, E. japonicus, Rosmarinusofficinalis L., andGenista monspessulana (L.) L.A.S. Johnson]. Twelve primary parasitoid species andtwo hyperparasitoids were reared; the most common were the egg parasitoid Trichogramma fasciatum(Perkins), the larval parasitoidsMeteorus ictericusNees, and Enytus eureka (Ashmead), and the pupalparasitoid Pediobius ni Peck.Meteorus ictericus accounted for �80% of the larval parasitoids, and wasrecovered from larvae collected on 39 plant species. Across all samples, mean parasitism was 84.4%for eggs, 43.6% for larvae, and 57.5% for pupae. The results are discussed with respect to the potentialfor resident parasitoid species to suppress E. postvittana populations.
KEY WORDS Lepidoptera, Totricidae, invasive species, host plant use, biological control
The light brown apple moth, Epiphyas postvittana(Walker), is a tortricid leafroller that is native tosoutheastern Australia (Danthanarayana 1975). It hasinvaded and established in western Australia, NewZealand, Caledonia, HI, the United Kingdom (Suck-ling and Brockerhoff 2010), and, most recently, Cal-ifornia (Brown et al. 2010). The moth is thought tohave Australian herbaceous plants as its ancestralhosts (Danthanarayana et al. 1995), but is known tofeed on numerous herbaceous plants, ornamentalshrubs and trees, and fruit and vegetable crops (Dan-thanarayana 1975, Geier and Briese 1980, Wearing etal. 1991). The long list of recorded host plants includesover 120 species found in Australia, including 23 Aus-tralian natives (Danthanarayana 1975, Geier and Bri-ese 1980), and 250 plant species found in New Zealand(Thomas and Shaw 1982). Epiphyas postvittana mayattack over 500 plant species in 363 genera (Sucklingand Brockerhoff 2010), and it has become a pest ongrapes, apples, caneberries, stone fruits, citrus, andnumerous ornamentals in Australia and New Zealand,and is considered to be an important invasive pest forNorth America and elsewhere (Varela et al. 2008,Brown et al. 2010, Suckling and Brockerhoff 2010).
The arrival of E. postvittana in California Þrst wasdocumented in 2006 when the moth was found inBerkeley, CA (Alameda Co.) (Brown et al. 2010). Thepest since has been found in 18 Californian counties,with the largest numbers of moths captured in theCentral Coast region (Monterey and Santa Cruz coun-ties) and Bay Area (which includes San Francisco,Alameda, and Contra Costa counties) (CDFA 2011).Currently, it is most often found on ornamental plantspecies in urban areas, with small incursions into someagricultural crops in the Central Coast (CDFA 2011).Commonly infested plants include species from di-verse geographic origins, such as a native Californianmanzanita, Arctostaphylos densiflora M.S. Baker; theMexican orange, Choisya dumosa (Torr.) A. Gray; aMediterranean native myrtle, Myrtus communis L.;Japanese cheesewood, Pittosporum tobira (Thunb.)W.T. Aiton; and Australian tea tree, Leptospermumlaevigatum (Sol. ex Gaertn.) F. Muell. (Mills et al.2009, Buergi et al. 2011). Although the availability ofsusceptible host plant species may not limit this pestÕsmovement in California, climate may prove to be thegreatest delimiting factor. Epiphyas postvittana favorsregions with mild summers (Thomson et al. 2010) andin California the pest has not yet been found in thewarmer interior valleys, where the hot summers maylimit its distribution (Gutierrez et al. 2010) into these1 Corresponding author, e-mail: [email protected].
0046-225X/12/0081Ð0090$04.00/0 � 2012 Entomological Society of America
agriculturally-rich regions. Because E. postvittanadoes not enter a winter diapause and has only a mod-erate cold hardness, cold winter temperatures alsomay limit its distribution further north in Californiaand North America (Burgi and Mills 2010).
Even with these possible climatic restrictions to itsmovement, E. postvittana has the potential to causeeconomic losses to CaliforniaÕs agriculture, nursery,and landscape systemÑincluding possible trade re-strictions (Varela et al. 2008). For these reasons, sys-tematic efforts were undertaken to eradicate this pestuntil spring 2010, when the program shifted to con-tainment rather than eradication. In the event that thispest could not be eradicated, we began investigatingthe potential for classical biological control and, toinitiate this work, we studied the impact of Californiaresident natural enemies onE. postvittana populationsin the Bay Area region. Prior Þeld surveys in SantaCruz and San Francisco counties recovered numerousparasitoid species from E. postvittana larvae and pu-pae, including Meteorus ictericus Nees and Enytus eu-reka (Ashmead) (Mills et al. 2009), and the egg para-sitoids Trichogramma fasciatum (Perkins) andTrichogramma platneri Nagarkatti (Roltsch et al.2009). However, the interactions among host plant,moth, and natural enemies, have yet to be determinedfor E. postvittana populations in California, where themothÕs potential host range is estimated to be over 150horticultural and 50 agricultural plants (CDFA 2011).
The aims of this study were to determine host plantuse by E. postvittana populations in the Bay Area, aswell as the species of indigenous parasitoids attackingthis invasive pest. The survey was conducted in theÔEast BayÕ of the San Francisco Bay Area., where E.postvittana was Þrst discovered and where there is agreat diversity of host plant ßora available. SpeciÞ-cally, we conducted a survey of plant species within adiscrete urban community to determine the occur-rence and relative abundance of E. postvittana andassociated parasitoids on different plant species. Wealso conducted general surveys, at four locations, onknown host plant species to determine parasitoid spe-cies composition and abundance. The results are dis-cussed with respect to the potential for resident para-sitoid species to suppressE. postvittanapopulations, aswell as eradication efforts given the pestÕs current hostrange.
Materials and Methods
Surveys were made in urban landscapes locatedwithin the city limits of Alameda, Albany, and Berke-ley (Alameda Co., CA); all in the East Bay.Complete Survey.A survey of 152 plant species was
conducted in the University of CaliforniaÕs residentialvillage in Albany (hereafter UC village). The UC vil-lage is a 23-ha student community, landscaped with awide variety of plant fauna that surrounds 75 apart-ment complexes, as well as parking facilities, openspaces, and driveways. More than 200 plant species aremanaged by either the groundÕs crew or residents,resulting in a diversity that includes annuals, biennials,
and perennials; shrubs, vines, fruit, and ornamentaltrees; potted vegetables and herbaceous plants; andground covers and weeds. Plant species were identi-Þed based on two garden plant books (Brenzel 2001,Brickell and Cathey 2004), and then veriÞed by takingphotographs that later were compared with the land-scape mapping provided by UC villageÕs maintenanceofÞce. Personnel from UC village groundÕs crewhelped conÞrm plant species identiÞcation. The sur-vey was conducted during September 2010, a periodcoinciding with higher larvalE. postvittana populationdensity during the second generation (Buergi et al.2011).
Representatives of all ornamental or horticulturalplant and weed species that were thought to supportlepidopteran larvae were inspected. For each plantspecies, a patch of land containing at least Þve indi-vidual plants was selected, although for some rarespecies fewer than Þve plants were sampled. For eachlarge plant, a 0.25-m2 section was randomly selected,with the sample area deÞned by a PVC pipe frame (0.5by 0.5 m). For each plant smaller than 0.25 m2, thewhole plant or patch was inspected, the sampled areawas measured, and data were transformed (insects perm2) for comparison among plant species. A singlepatch of Þve plants was sampled for most of the 152plants species inspected, the exceptions being rareplant species where a patch of Þve plants could not befound.
The foliage within the sample region was thor-oughly inspected for eggs, larvae, and pupae of E.postvittana. Other lepidopteran species were noticedoccasionally but were not recorded. All observed E.postvittana and parasitoid cocoons were collected andbrought to the laboratory where egg masses, pupae,and parasitoid cocoons were reared individually invials (1 by 5 cm). Larvae were reared individually onbouquets of French broom, isolated in plastic con-tainers (150 ml) and held under controlled conditions(24 � 2�C, 40Ð60% RH), with fresh bouquets added asneeded. Material was held until the emergence ofadult moths or parasitoids; all dead larvae and pupaewere dissected for the presence or absence of para-sitoids.General Survey. Additional surveys were con-
ducted at four locations on Þve plant species known tobe commonly infested by E. postvittana.The locationsand plant species were 1) UC village for myrtle androsemary, Rosmarinus officinalis L.; 2) the Universityof CaliforniaÕs Lawrence Hall of Science (hereafterUC Lawrence), located in the Berkeley hills, forFrench broom, Genista monspessulana (L.) L.A.S.Johnson; 3) the University of CaliforniaÕs Oxford TractFacility (hereafter Oxford Tract), located northwestof the campus, for Japanese spindletree; and 4) anapartment complex in Alameda, for Japanese cheese-wood. For these general surveys, plots were composedof 10Ð20 sampled plants that were located in areas thatranged in size from 5.8 to 19.8 m2, except the UCLawrence site where plants were randomly sampledfrom patches of French broom grown around parkinglots. Surveys were conducted from June or July to
82 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 1
October 2010, with samples taken approximately every2 wk at each site. On each sample date a 0.25-m2 plantsectionwassearchedandallobservedE.postvittanawerecollected, brought to the laboratory, and reared to adultmoths or parasitoids, as described previously.Moth Identification. Voucher specimens of adult
moths reared are in the UC Berkeley Essig Museum.During the project, to verify the correct identiÞcationof E. postvittana, DNA extractions, using the DNeasytissue kit (Qiagen, Inc., Valencia, CA), were per-formed on subsamples collected from 40 of the sam-pled plant species. A segment of the CO1 gene wasampliÞed with primers described by Hebert et al.(2003). An initial denaturing step at 95�C for 5 min wasfollowed by 33 cycles of 30 s at 94�C, 1 min 30 s at 51�C,and 1 min at 72�C; with a Þnal extension of 5 min at72�C. All reactions usedTaqpolymerase chain reaction(PCR) Master Mix Kit (Qiagen) with a Þnal MgCl2concentration of 1.5 mM and 0.125 �M of each primer.A 0.5-�l aliquot of genomic DNA was used per reac-tion in a total reaction volume of 12.5 �l. PCR wasperformed in a Biometra T- personal thermal cycler(Biometra, Horsham, PA) and products were visual-ized after electrophoresis on 1.2% agarose gels, stainedwith ethidium bromide. PCR products then werecleaned using the QIAquick PCR PuriÞcation Kit(Qiagen) and submitted to the UC Berkeley DNAsequencing facility for direct sequencing in both di-rections using the ABI Big Dye V3.1 terminator se-quencing reaction kit (Perkin-Elmer) on an ABI3707xl DNA Analyzer (Perkin-Elmer) with POP sevenand a 50-cm array. DNA sequence results were in-spected and aligned manually in SeqMan two version5.07 (DNASTAR, Madison, WI) and the identity of thesamples was conÞrmed using the Basic Local Align-ment Search Tool (BLAST) on GenBank. All pro-cessed samples matched E. postvittana haplotypespublished by Tooman et al. (2011), showing 99Ð100%similarity.DataAnalysis.Resultsarepresentedasmeans(�SE).
Average parasitism and the percentage of major parasi-toid species reared were compared among different lo-cations. Before data analysis, proportional data were arc-sine square-root transformed to satisfy the assumptionsof analysis of variance (ANOVA). Data were analyzedusing one-way ANOVA for comparisons of means andTukeyÕs HSD test for multiple comparisons of means(JMP software, V. 8.0.3, SAS 2008, Cary, NC).
Results
Host Plant Use. During the complete survey, 152plant species (66 families) were inspected at the UCvillage, representing a diverse assemblage of taxonom-ically dissimilar species (Fig. 1). Epiphyas postvittanawas found on 75 species belonging to 36 families and,of these, 34 species from 27 families appear to be newhost records (Table 1). Hosts included shrubs (39);ground covers (herbs, ßowers, and weeds) (26); or-namental and fruit trees (8); and climbing vines (2).Of those plant species with a clearly known nativeorigin (as described by Brenzel 2001, Brickell and
Cathey 2004), 77.8% of the Mediterranean plants wereinfested, 47.8% of the Asian plants, 47.5% of the West-ern Hemisphere plants (including North, Central, andSouth Americas), 69.3% of the European (excludingMediterranean) plants, and 40.0% of the African plantssampled in the survey.
Of the nine plants surveyed considered to be Cal-ifornia native species, two (22.2%) were infested: Lit-tle Sur Manzanita,Arctostaphylos edmundsii J. T. How-ell, and tree anemone, Carpenteria californica Torr. Incomparison, of the nine Australian native plants sur-veyed, Þve (55.6%) were infested. Overall, plant spe-cies in the Rosaceae (14) and Fabaceae (7) were themost common E. postvittana hosts; however, in mostplant families surveyed there were some representa-tive species infested (Fig. 1).Epiphyas postvittana larvae were found on 70 of 75
infested plant species, whereas the remaining Þve plantspecies had only egg or pupal stages (Table 1). Eggs ofE. postvittanawere not easily found, with the exceptionof samples on Pittosporum species, and few pupae (�0.2per m2) were found in comparison to larvae (�5.0 perm2), in part because of the seasonal timing of the survey.For this reason, only the relative abundance of larvae ispresented (Table 1). The heavily infested plants in-cluded weeds [wild mint, Mentha arvensis L., purpleglory tree, Tibouchina semidecandra (Schrank & Mart.ex. DC.) Cogn., horseweed, Pelargonium sp., and com-mon ßax, Linum usitatissimum L.]; ornamental shrubs(tree anemone, Japanese spindletree, myrtle, bluebellcreeper, Sollya heterophylla Lindl.); and weedy shrubs(French broom and blue leadwood, Ceratostigmaplumbaginoides Bunge) (Table 1). The highest infesta-tion level was on myrtle (37.6 larvae per m2), where E.postvittana feeding resulted in the dieback of foliage onsome of the sampled plants.
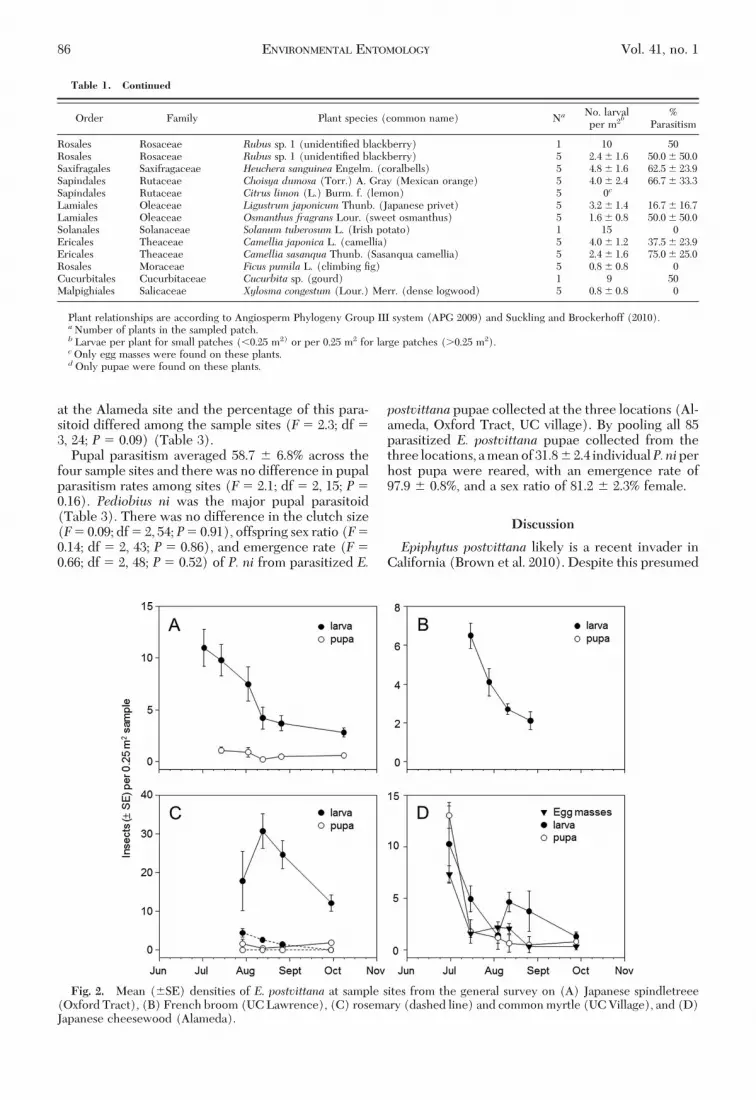
Fromthegeneral survey,E.postvittana larvaldensitieson the Þve surveyed plant species show a decline inpopulation density at all sites during the sampling period(Fig. 2). As before, egg masses and pupae were moredifÞcult toÞnd.EggswereeasilyvisibleonlyonJapanesecheesewood plants. Pupae were not found at the UCLawrence site. At the Alameda site, whereE. postvittanaeggmasses, larvae,andpupaewereall foundonJapanesecheesewood, there was seasonal decline (June to Octo-ber) of all development stages (Fig. 2d).Parasitoid Species. In total, twelve primary parasi-
toid species were found attacking E. postvittana eggs,larvae, and pupae from the complete and general sur-veys (Table 2). From the complete survey at the UCvillage, parasitism ranged from 0 to 100% per infestedplant species (Table 1). Only six E. postvittana eggmasses were found at the UC Village and all (100%)were parasitized by either Trichogramma fasciatum(Perkins) orTrichogrammaplatneriNagarkatti. Larvalparasitism averaged 35.5 � 2.9% (n � 157), across allplant species. Two solitary larval endoparasitoids,Me-teorus ictericus Nees and Enytus eureka (Ashmead),accounted for 76.9% and 16.7% of the larval parasitism,respectively. A single, unidentiÞed Apanteles sp. alsowas reared, however its rarity suggests this species didnot commonly attackE. postvittanaor was reared from
February 2012 WANG ET AL.: LIGHT BROWN APPLE MOTH HOST PLANTS AND PARASITOIDS IN CALIFORNIA 83
a different tortricid species that was inadvertentlycollected during the samples. Meteorus ictericus wasfound on 39 of 70 E. postvittana larval-infested plantspecies, whereasE. eurekawas found only on 10 of thesurveyed plant species. Four solitary larvalÐpupalendo-parasitoids species were recovered, accountingfor the remaining 6.4% larval parasitism. These werethe ichneumonids Agrypon clandestinum (Graven-horst) and Exochus nigripalpus Thompson, and thetachinids Actia interrupta Curran and Nemorilla pyste(Walker) (Table 2). Pupal parasitism was 33.7 � 7.4%(n� 37) averaged across all samples at the UC village;a gregarious endo-parasitoid,PediobiusniPeck (Hym.:Chalcididae) accounted for all pupal parasitism. Twohyperparasitoid species were also reared, Dibrachyscavus (Walker) (Hym.: Pteromalidae) and Conuraside (Walker) (Hym.: Chalcididae), both from co-coons of the primary parasitoid E. eureka.
In the complete survey (UC village), percentageparasitism was not related to E. postvittana larval den-sity per plant (linear regression: F� 0.072; df � 1, 259;P � 0.788; r2 � 0.0003).
During the general surveys (all four sample sites),in addition to the parasitoids described previously, the
ichneumonid larvalÐpupal parasitoid Ischnus inquisi-torius (Muller) and the pupal parasitoid Brachymeriaovata (Say) were recovered (Table 2) at the OxfordTract, UC Lawrence, and Oxford Tract sites. As men-tioned previously, E. postvittana eggs rarely werefound during the general surveys, except on Japanesecheesewood at the Alameda site, where a mean of5.8 � 2.4 egg masses per sample date (n � 6) werecollected, of which 84.4 � 7.5% were parasitized byeither T. fasciatum (95%) or T. platneri (5%). Larvalparasitism averaged 43.9 � 4.7% across the four samplesites, and parasitism rates were similar at the OxfordTract (45.1%), UC village (45.9%) and Alameda(53.6%) sites, which were higher than the UC Law-rence (19.5%) site (F � 4.2; df � 3, 24; P � 0.016)(Table 3). There were fewer larval parasitoid speciescollected at the UC Lawrence site, but lower larvalparasitism at this site resulted from lower levels byM.ictericus and higher levels by E. eureka, in comparisonwith the other three sites (Table 3).Enytus eurekawasthe dominant parasitoid at the UC Lawrence site (F�6.3; df � 3, 24; P� 0.05), whereasM. ictericuswas thedominant parasitoid at the other three sites (F� 14.2;df � 3, 24; P� 0.001). No A. clandestinumwere found
Fig. 1. Interrelationships of surveyed plant species (orders and families) and host record frequency for E. postvittana inUC Village, Albany, CA; after each family name the number of plant species inspected and infested (in the parentheses) byE. postvittana are provided. Plant relationships are according to Angiosperm Phylogeny Group III system (APG III 2009) andSuckling and Brockerhoff (2010).
84 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 1
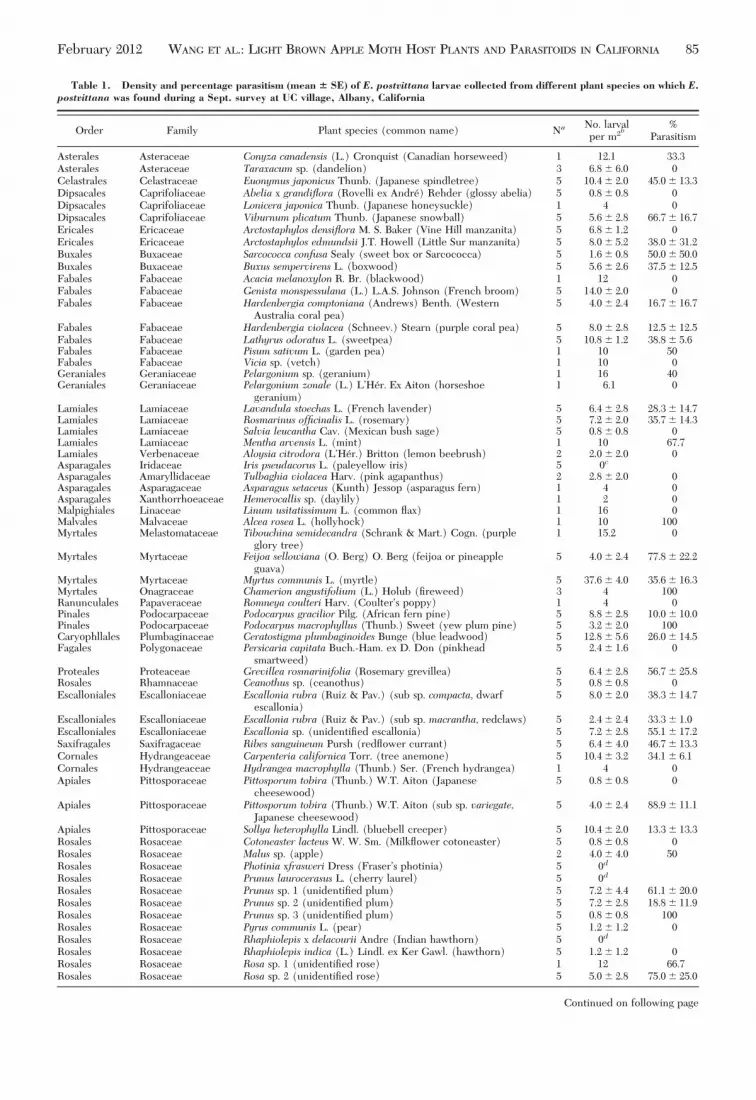
Table 1. Density and percentage parasitism (mean � SE) of E. postvittana larvae collected from different plant species on which E.postvittana was found during a Sept. survey at UC village, Albany, California
Order Family Plant species (common name) NaNo. larvalper m2b
Rosales Rosaceae Rhaphiolepis indica (L.) Lindl. ex Ker Gawl. (hawthorn) 5 1.2 � 1.2 0Rosales Rosaceae Rosa sp. 1 (unidentiÞed rose) 1 12 66.7Rosales Rosaceae Rosa sp. 2 (unidentiÞed rose) 5 5.0 � 2.8 75.0 � 25.0
Continued on following page
February 2012 WANG ET AL.: LIGHT BROWN APPLE MOTH HOST PLANTS AND PARASITOIDS IN CALIFORNIA 85
at the Alameda site and the percentage of this para-sitoid differed among the sample sites (F � 2.3; df �3, 24; P � 0.09) (Table 3).
Pupal parasitism averaged 58.7 � 6.8% across thefour sample sites and there was no difference in pupalparasitism rates among sites (F � 2.1; df � 2, 15; P �0.16). Pediobius ni was the major pupal parasitoid(Table 3). There was no difference in the clutch size(F� 0.09; df � 2, 54; P� 0.91), offspring sex ratio (F�0.14; df � 2, 43; P � 0.86), and emergence rate (F �0.66; df � 2, 48; P � 0.52) of P. ni from parasitized E.
postvittana pupae collected at the three locations (Al-ameda, Oxford Tract, UC village). By pooling all 85parasitized E. postvittana pupae collected from thethree locations, a mean of 31.8 � 2.4 individualP. niperhost pupa were reared, with an emergence rate of97.9 � 0.8%, and a sex ratio of 81.2 � 2.3% female.
Discussion
Epiphytus postvittana likely is a recent invader inCalifornia (Brown et al. 2010). Despite this presumed
Table 1. Continued
Order Family Plant species (common name) NaNo. larvalper m2b
Plant relationships are according to Angiosperm Phylogeny Group III system (APG 2009) and Suckling and Brockerhoff (2010).aNumber of plants in the sampled patch.b Larvae per plant for small patches (�0.25 m2) or per 0.25 m2 for large patches (�0.25 m2).cOnly egg masses were found on these plants.dOnly pupae were found on these plants.
Fig. 2. Mean (�SE) densities of E. postvittana at sample sites from the general survey on (A) Japanese spindletreee(Oxford Tract), (B) French broom (UC Lawrence), (C) rosemary (dashed line) and common myrtle (UC Village), and (D)Japanese cheesewood (Alameda).
86 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 1
short history, the current survey shows that E. post-vittana has established on a wide diversity of plantspecies in the urban landscape of the San FranciscoBay Area. Most of the used host plants were not Aus-tralian natives, which were imported primarily forlandscaping, but rather came from diverse temperateand subtropical regions. Host plants also included Cal-ifornia native plants. As has been noted, E. postvittanademonstrates a remarkable adaptation to various hostplant species (Danthanarayana 1975) that may aid inits exceptional ability to expand its host range in exoticenvironments (Suckling and Brockerhoff 2010). Cal-iforniaÕs urban landscaping provides a diversity of na-tiveandexoticplant species,which this survey showedto contain a suite of acceptable food sources for thisinvasive species. Although urban areas are a target ofongoing containment efforts (e.g., insecticides, sterilemale releases), the vast number and diversity of suit-able host plants reduces the feasibility of local erad-ication within CaliforniaÕs urban environments. More
importantly, survey results suggest that urban areasmay provide a refuge for this pest, even if suppressionprograms in outlying agricultural areas are successful.
The plant species survey at the UC village waslimited to a single residential community in the EastBay, and our results most likely underestimated therange of plant species suitable for attack because sam-ples were taken in September, when we expectedhigher larval E. postvittana densities (Buergi et al.2011), but when in fact the seasonal population den-sity already was in decline (Fig. 2). The observedreduction in collectedE. postvittana in September andOctober may have been caused, in part, by emergenceof adult moths, which were not sampled. Therefore,the survey provides a conservative estimate of thediversity of host plants used by E. postvittana. Never-theless, we found 75 plant species used by E. postvit-tana in this one urban setting. There is not a completelist of E. postvittana host plants to which we can com-pare our survey results. Many reported hosts have not
Table 2. Primary parasitoid species collected from LBAM in the east Bay Area, California
Voucher specimens have been placed in the University of California Insectary and Quarantine collection.
Table 3. Hosts sampled, host stage sampled, number of samples (N), parasitoid species reared, and percentage parasitism of E.postvittana from June to September 2010 at four different locations in the Bay Area during the general survey
aData from two plants (R. officinalis and M. communis) in the UC village were pooled.
February 2012 WANG ET AL.: LIGHT BROWN APPLE MOTH HOST PLANTS AND PARASITOIDS IN CALIFORNIA 87
been identiÞed to species and, of those plant specieslisted, there may be a bias toward those of economicimportance (Suckling and Brockerhoff 2010). A re-cent review listed 135 genera or species that could hostE. postvittana, and although this was described as apartial list, most host species were in the Rosaceae(13), Asteraceae (12), and Fabaceae (11) (Brown etal. 2010). Similarly,we foundmost speciesofhostplantto be in the Rosaceae and Fabaceae (Table 1). Still, italso was common that plant families represented byonly one or two species in our surveys also wereinfested (Table 1; Fig. 1).
Most of the collected E. postvittana larvae werethird to Þfth instars, indicating successful develop-ment of the mothÕs larvae on these host plants. Weobserved that infested plant species were character-ized by soft or Þne leaf texture and dense foliage, suchas Irish potato (Solanum tuberosum L.), common ßax,vetch (Vicia spp.), glorytree (Tibouchina spp.), horse-weed, Japanese cheesewood, and bluebell creeper(Table 1). For some plant species only the fresh grow-ing foliage was infested, such as the Japanese privet(Ligustrum japonicum Thunb.) and sweet osmanthus(Osmanthus fragrans Lour.). Plants with dense orwhorled leaves also were commonly infested, such asFrench broom, leadwort (Plumbago spp.), and myrtle;as were plant species with soft needle-like leaves, suchas yew plum pine [Podocarpusmacrophyllus (Thunb.)Sweet], fern pine (Podocarpus graciliorPilger), scarletsprite (Grevillea rosmarinifolia A. Cunn.), and aspar-agus fern [Asparagus setaceus (Kunth) Jessop] (Table1). In contrast, plants with little new foliage growthduring the survey (e.g., hardened leaves) rarely wereinfested, such as oaks (Quercus spp.), CA buckeye[Aesculus californica (Spach) Nutt], magnolia (Mag-nolia spp.), loquat [Eriobotrya japonica (Thunb.)Lindl.], and wavyleaf silktasse (Garrya ellipticaDoug-las ex Lindl.) (Table 1). Besides plant texture andstructure, E. postvittana performance and host pref-erences also may be affected by plant physiological orchemical factors (Rojas et al. 2003, Pontes et al. 2010).
For many of the annual ground covers, high levelsof infestation found in our September surveys may notbe indicative of the plantsÕ year-round importance.For example, whereas many of the Fabaceae appear tosupport high E. postvittana densities, some of theseplant species are not available year-round or may notbe suitable throughout the season. Most of the patchessampled had clearly deÞned borders with other spe-cies, reducing the possibility that larvae moved fromanother host onto the sampled plant, but this move-ment also is a possibility.
A rich diversity of resident parasitoid species wasfound attackingE. postvittana in the San Francisco BayArea. Few egg masses were collected in our surveys,but those collected were often parasitized by eitherTrichogramma fasciatum or T. platneri. Most E. post-vittana eggs were collected on Japanese cheesewoodat the Alameda site from late June to early July, withT. fasciatum commonly reared and T. platneri rarelyreared. Egg parasitoids were reared previously fromsentinel eggs of E. postvittana deployed in Santa Cruz
and San Francisco, but the Santa Cruz site primarilyconsisted ofT. platneri in the spring and early summer,and the San Francisco collections were dominated byT. fasciatum in the fall (Roltsch et al. 2009). In Aus-tralia and New Zealand, a number of Trichogrammaspecies are associated with E. postvittana (Paull andAustin 2006, Suckling and Brockerhoff 2010), and astudy of Trichogramma funiculatumCarver, in Austra-lia, suggested that egg parasitism similarly was higherin late summer or fall (Danthanarayana 1980).
From both surveys, Meteorus ictericus, Enytus eu-reka, and Agrypon clandestinum were the three mostcommonly reared larval parasitoid species, althoughM. ictericuswas clearly the more dominant of the threespecies (Table 3). Other parasitoids included the ta-chinidsActia interrupta andNemorilla pyste,which arerelatively common parasitoids of Nearctic tortricids(OÕHara 2005). Two pupal parasitoids were found,withPediobius ni accounting for 30.0Ð78.1% parasitismof collected pupae, although the number of pupaecollected was low. The diversity of pupal parasitoids inour survey may be under-represented as relatively fewpupae were recovered. In its native Australian range(Paull and Austin 2006) and introduced New Zealandrange (Thomas 1989), numerous parasitoid specieshave been reared fromE.postvittana larvae and pupae.Key parasitoid species include larval parasitoidsDolichogenidea tasmanica (Cameron) and Goniozusjacintae Farrugia, the tachinid ßy Trigonospila brevi-facies (Hardy), and the pupal parasitoidXanthopimplarhopaloceros Krieger (Suckling and Brockerhoff2010). Biological control has been demonstrated to becapable of suppressing E. postvittana in New Zealand,where the pest has been successfully managedthrough a combination of biological control andthreshold-based applications of selective insecticide(Varela et al. 2010).
The generalist parasitoids reared in our surveylikely moved over from tortricid or related speciesalready present in California. For example, an earliersurvey reared numerous ichneumonoids from Califor-nia tortricids such as Acleris variegana (Schiffer-muller) (garden rose tortrix), Croesia albicomana(Clemens), Archips argyrospilus (Walker) (fruit treeleafroller), Argyrotaenia franciscana (Walsingham)(orange tortrix), and Pandemis pyrusana (Kearfott)(apple pandemis) (Powell 1962). Other westernNorth American leafroller species with relatively largecomplexes of natural enemies that might attack E.postvittana include Spilonota ocellana (Denis andSchiffermuller) (eye-spotted bud moth), Ancyliscomptana(Froelich) (strawberry leafroller), andCho-ristoneura rosaceana (Harris) (obliquebanded leafrol-ler) (Obrycki et al. 1993, Krugner et al. 2007, Fernan-dez-Triana and Huber 2010). For example, of the mostcommon parasitoids reared, M. ictericus has been re-corded from S. ocellana in California (Madsen andBorden 1949) andE. eurekahas been recorded fromA.variegana in California (N.J.M., unpublished observa-tions) and C. rosaceana in North America (Coop et al.1989).
88 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 1
Here, we assume that all parasitoids in our surveyswere reared from E. postvittana rather than from anyother tortricid species, although we did not make anidentiÞcation of the remains of leafroller eggs, larvae,or pupae from which the parasitoids were reared.Molecular identiÞcation of the subsampled specimensshowed onlyE. postvittana, and the reared adult mothswere identiÞed morphologically as E. postvittana. Inaddition, the majority of the other tortricid speciesexpected to be found on the sampled plants are uni-voltine and would not be found as larvae during theprimary collection period (September). The excep-tions are A. franciscana and Clepsis peritana (Clem-ens) (garden tortrix), both of which are smaller tor-tricid species, and in the case of A. franciscanasupporting a very different assemblage of parasitoidspecies (Coop et al. 1989, Walker and Welter 2004)that were not reared in either of our surveys.
Many factors may inßuence patterns in parasitoidassemblages. In general, host characteristics (such ashost plant range and abundance) and habitat charac-teristics (such as habitat diversity and stability) affectthe diversity of parasitoids (Hawkins and Sheehan1994). There is often a positive relationship betweenlandscape vegetation complexity and natural enemyabundance and diversity (Marino et al. 2006). In thecase of E. postvittana, the parasitoid assemblage wascomposed of generalist species. The heavily land-scaped urban environment may be an ideal habitat forgeneralist parasitoids because it provides a great di-versity of host plants and microclimates, which alsomay result in numerous lepidopteran host species.Although the species richness of parasitoids was rel-atively high in our survey, the evenness among para-sitoid species was not. We had expected that the di-versity of host plants used byE. postvittana in our Þeldsites would have resulted in both a richness and even-ness of parasitoid species that could provide betterpest suppression (Crowder et al. 2010). Instead, therewas commonly dominance by a single parasitoid spe-cies at each site surveyed. For example, M. ictericuswas the dominant larval parasitoid at the Oxford Tract,Alameda, and UC village, but at the UC Lawrence siteE. eureka and A. clandestinum were more common(Table 2). The UC Lawrence site, in the Berkeley hills,differs from the other three sites in that it is morenaturally landscaped and has less plant diversity, beingprimarily French broom and grasses in the under sto-rey of a eucalyptus forest.
Our results show that resident parasitoids may becontributing to the suppression of E. postvittana pop-ulations in California, at least on the host plant speciesmonitored in our surveys. Further work may showwhether adequate suppression ofE. postvittana can beachieved by resident parasitoids present in the urbanenvironments of California or whether introductionsof specialized parasitoid species from Australia wouldaid in the future management of this new invader. Wesuggest that as E. postvittana expands its range inCalifornia, additional parasitoid species will be foundusing this pest as a novel host. Further studies willbetter elucidate the levels of pest suppression by abi-
otic and biotic factors in California and help to de-termine the need for classical biological controls, aswell as targeting those natural enemies that might bestÞt into the parasitoid assemblage already present inCalifornia. As discussed here, these relationships maybe governed, in part, by diversity of host plants avail-able for E. postvittana and other leafrollers, as well asregional differences that inßuence temperature andrainfall (Gutierrez et al. 2010). The introduction ofexotic parasitoids from Australia is being consideredwith respect to the diversity of natural enemies al-ready present in California, the chance that any in-troduced parasitoid may interfere with resident para-sitoids, and the development of a strategic plan for thelong-term management of this pest in California.
Acknowledgments
We thank numerous taxonomists that helped identify theparasitoids, including John Luhman (University of Minne-sota), James OÕ Hara (Invertebrate Biodiversity, Agricultureand Agri-Food Canada), and John Heraty (UC Riverside).We also thank William Roltsch (California Department ofFood and Agriculture) for conÞrming the Trichogrammaspecies and facilitating Þeld studies; Alan Sawyer (USDA-APHIS-PPQ-CPHST) for facilitation of the research;Mathew Middleton (UC Berkeley) for molecular analysis;Addie Abrams and Elizabeth Pierson (UC Berkeley) for Þeldand laboratory assistance; Stephen Pree (UC village GroundSuperintendent) and Tina Wistrom (UC Berkeley) for iden-tifying plant species; and Brian Hogg and Linda Burgi (UCBerkeley) for help and useful suggestions on this study.Funding was supported by USDA-APHIS08-8100-1310.
References Cited
APG III. 2009. On update of Angiosperm Phylogeny GroupclassiÞcation for the orders and families of ßoweringplants: APG III. Bot. J. Linn. Soc. 161: 105Ð121.
Brenzel, K. N. 2001. Western garden book. Sunset Publish-ing Corporation, Menlo Park, CA.
Brickell, C., and H. M. Cathey. 2004. A-Z encyclopedia ofgarden plants. DK Publishing, Inc, New York.
Brown, J. W., M. E. Epstein, T. M. Gilligan, S. C. Passoa, andJ. A. Powell. 2010. Biology, identiÞcation, and history ofthe light brown apple moth, Epiphyas postvittana(Walker) (Lepidoptera: Tortricidae: Archipini). Am. En-tomol. 56: 34Ð43.
Burgi, L. P., and N. J. Mills. 2010. Cold tolerance of theoverwintering larval instars of light brown apple mothEpiphyas postvittana. J. Insect Physiol. 56: 1645Ð1650.
Buergi, L. P., W. J. Roltsch, and N. J. Mills. 2011. Abundance,age structure, and voltinism of light brown apple moth pop-ulations in California. Environ. Entomol. 40: 1370Ð1377.
[CDFA] California Department of Food and Agriculture.2011. Light brown apple moth project. (http://www.cdfa.ca.gov/phops/PDEP/lbam_main.htm/).
Coop, L. B., A. Knight, and G. Fisher. 1989. Parasitism oforange tortrix on caneberry, Rubus spp. in western Or-egon and Washington. J. Entomol. Soc. B.C. 86: 63Ð65.
Crowder, D. W., T. D. Northfield, M. R. Strand, and W. E.Snyder. 2010. Organic agriculture promotes evennessand natural pest control. Nature 466: 109Ð112.
Danthanarayana, W. 1975. Bionomics, distribution and hostrange of light brown apple moth, Epiphyas postvittana(Walker) (Tortricidae). Aust. J. Zool. 23: 419Ð437.
February 2012 WANG ET AL.: LIGHT BROWN APPLE MOTH HOST PLANTS AND PARASITOIDS IN CALIFORNIA 89
Danthanarayana, W. 1980. Occurrence of Trichogrammafuniculatum, an egg parasitoid of the light brown applemoth, Epiphyas postvittana. Entomol. Exp. Appl. 28: 287Ð294.
Danthanarayana, W., H. Gu, and S. Ashley. 1995. Popula-tion growth potential of Epiphyas postvittana, the lightbrown apple moth (Lepidoptera: Tortricidae), in relationto diet, temperature and climate. Aust. J. Zool. 43: 381Ð394.
Fernandez-Triana, J. L., and J. T. Huber. 2010. Braconidparasitoids (Hymenoptera: Braconidae) of NearcticCho-ristoneura species (Lepidoptera: Tortricidae), with asummary of other parasitoid families attacking Choristo-neura. Can. Entomol. 142: 295Ð343.
Geier, P. W., and D. T. Briese. 1980. The light brown applemoth, Epiphyas postvittana (walker). 4. Studies on pop-ulation dynamics and injuriousness to apples in the Aus-tralian Capital Territory. Aust. J. Ecol. 5: 63Ð93.
Gutierrez, A. P., N. J.Mills, and L. Ponti. 2010. Limits to thepotential distribution of light brown apple moth in Ari-zona-California based on climate suitability and hostplant availability. Biol. Invasions 12: 3319Ð3331.
Hawkins, B. A., and W. Sheehan. 1994. Parasitoid commu-nity ecology. Oxford University Press, Oxford, UnitedKingdom.
Hebert, P.D.N., A. Cywinska, S. L. Ball, and J. R. deWaard.2003. Biological identiÞcations through DNA barcodes.Proc. R. Soc. Biol. Sci. Ser. B. 270: 313Ð321.
Krugner, R., K. M. Daane, A. B. Lawson, and G. Y. Yokota.2007. Temperature-dependent development of Macro-centrus iridescens (Hymenoptera: Braconidae) as a para-sitoid of the obliquebanded leafroller (Lepidoptera: Tor-tricidae): implications for Þeld synchrony of parasitoidand host. Biol. Control 42: 110Ð118.
Madsen, H. F., and A. D., Borden. 1949. The eye-spottedbud moth on prune in California. J. Econ. Entomol. 42:915Ð920.
Marino, P. C., D. A. Landis, and B. A. Hawkins. 2006. Con-serving parasitoidassemblagesofNorthAmericanpestLep-idoptera: does biological control by native parasitoids de-pend on landscape complexity? Biol. Control 37: 173Ð185.
Mills, N., L. Burgi, and W. Roltsch. 2009. Seasonal patternsof activity and larval parasitism of light brown apple mothin two coastal areas of California, pp. 12Ð14. In D. M.Woods (ed.), Biological Control Program 2008 AnnualSummary. California Department of Food and Agricul-ture, Plant Health and Pest Prevention Services, Sacra-mento, CA.
O’Hara, J. E. 2005. A review of the tachinid parasitoids(Diptera: Tachinidae) of Nearctic Choristoneura species(Lepidoptera: Tortricidae), with keys to adults and pu-paria. Zootaxa: 1Ð46.
Obrycki, J. J., A. M. Ormord, A. D. Gabriel, and C. J. Orr.1993. Larval and pupal parasitism of the strawberry lea-froller (Lepidoptera: Tortricidae). Environ. Entomol. 22:679Ð683.
Paull, C., and A. D. Austin. 2006. The hymenopteran para-sitoids of light brown apple moth, Epiphyas postvittana(Walker) (Lepidoptera: Tortricidae) in Australia. Aust. J.Entomol. 45: 142Ð156.
Pontes,W.J.T., E. R. Lima, E. G. Cunha, P.M.T.DeAndrade,A. P. Lobo, and R. Barros. 2010. Physical and chemicalcues affect oviposition by Neoleucinodes elegantalis.Physiol. Entomol. 35: 134Ð139.
Powell, J. A. 1962. Host-parasite relationships of CaliforniaTortricinae. Pan-Pac. Entomol. 2: 131Ð140.
Rojas, J. C., A. Virgen, and L. Cruz-Lopez. 2003. Chemicaland tactile cues inßuencing oviposition of a generalistmoth, Spodoptera frugiperda (Lepidoptera: Noctuidae).Environ. Entomol. 32: 1386Ð1392.
Roltsch, W., N. Carruthers, R. Stouthamer, and N. Saechao.2009. Parasitism and predation of light brown apple motheggs, pp. 9Ð11. InD. M. Woods (ed.), Biological ControlProgram 2009 Annual Summary. California Departmentof Food and Agriculture, Plant Health and Pest Preven-tion Services, Sacramento, CA.
Suckling, D. M., and E. G. Brockerhoff. 2010. Invasion bi-ology, ecology, and management of the light brown applemoth (Tortricidae). Annu. Rev. Entomol. 55: 285Ð306.
Thomas, W. P. 1989. Epiphyas postvittana (Walker), lightbrown apple moth (Lepidoptera: Tortricidae), pp. 187Ð195. InP. J. Cameron, R. L. Hill, J. Bain, and W. P. Thomas(eds.),A reviewofbiological controlof invertebratepestsand weeds in New Zealand, 1874 to 1987. CAB Interna-tional, Wallingford, United Kingdom.
Thomas,W. P., and P.W. Shaw. 1982. An attempt to controlthe light brown apple moth, Epiphyas postvittana(Walker) (Lepidoptera: Tortricidae), by male removal,pp. 33Ð45. DSIR Entomology Division Report, DSIR,Auckland, NZ.
Thomson, L. J., S. Macfadyen, and A. A. Hoffmann. 2010.Predicting the effects of climate change on natural ene-mies of agricultural pests. Biol. Control 52: 296Ð306.
Tooman, L. K., C. J. Rose, C. Carraher, D. M. Suckling, S. R.Paquette, L. A. Ledezma, T.M.Gilligan,M.Epstein,N. B.Barr, and R. D. Newcomb. 2011. Patterns of mitochon-drial haplotype diversity in the invasive pest Epiphyaspostvittana (Lepidoptera: Tortricidae). J. Econ. Entomol.104: 920Ð932.
Varela, L. G., M. W. Johnson, L. Strand, C. A. Wilen, and C.Pickel. 2008. Light brown apple mothÕs arrival in Cali-fornia worries commodity groups. Calif. Agric. 62: 57Ð61.
Varela, L. G., J.T.S. Walker, P. L. Lo, and D. J. Rogers. 2010.New Zealand lessons may aid efforts to control lightbrown apple moth in California. Calif. Agric. 64: 6Ð12.
Walker, K. R., and S. C. Welter. 2004. Biological controlpotential of Apanteles aristoteliae (Hymenoptera: Bra-conidae) on populations of Argyrotaenia citrana (Lepi-doptera: Tortricidae) in California apple orchards. Envi-ron. Entomol. 33: 1327Ð1334.
Wearing, C. H., W. P. Thomas, J. S. Dugdale, and W. Dan-thanarayana. 1991. Tortricid pests of pome and stonefruits, Australian and New Zealand species, pp. 453Ð472.In L. van der Geest and H. H. Evenhuis (eds.), Tortricidpests: their biology, natural enemies and control. Elsevier,Amsterdam, the Netherlands.