jo ur nal home p ag e wwwelsev ier com locate dendro
RIGINAL ARTICLE
ariation in radial growth of Pinus cooperi in response to climaticignals across an elevational gradient
ariacuten Pompa-Garciacuteaalowast Juliaacuten Cerano-Paredesb Peter Z Fuleacutea
School of Forestry Northern Arizona University PO Box 15018 Flagstaff AZ 86011-5018 United StatesInstituto Nacional de Investigaciones Forestales Agriacutecolas y Pecuarias Centro Nacional de Investigacioacuten Disciplinaria en Relacioacuten Agua Suelo Plantatmoacutesfera Gomez Palacio Durango 35140 Mexico
r t i c l e i n f o
rticle historyeceived 10 March 2013ccepted 30 May 2013
eywordsree-rings
a b s t r a c t
Climate change could modify the biogeography of many forest species Elevational gradients have beendocumented as strategic sites to better understand tree growth response to regional climate variablesPinus cooperi Blanco is one of the most important species in Northern Mexico However little is knownconcerning effects of climate responses on growth of this species We used tree data records to compare theinfluence of precipitation and temperature on radial growth among P cooperi populations across a moun-
tain landscape at elevation gradient Correlation and regression analysis of the regional growthndashclimaterelationships showed that radial growth was correlated with previous winter conditions at most sitesalong the gradient Wet and cold winters were positively associated with radial growth Although ourresults showed significant climate influences on tree radial growth other site factors also may haveaffected growthndashclimate responses The results support the idea that climate change influences P cooperigrowth
udtcg2s(
bCl(di
ntroduction
Several studies have documented that forest ecosystems coulde at risk due to global climatic changes (Seager et al 2007 Huangt al 2010 Candel-Peacuterez et al 2012) Increasing temperatures arexpected by the 2050s and droughts are predicted to be moreevere than any in the last 100 years (Williams et al 2012) Its important to understand the repercussions of these climatichanges for the distribution and composition of vegetation Cli-atic changes may impact plant phenology growth rate forest
xtent and other gradual changes in population processes (Lo et al010) Understanding climatendashtree growth relationships is a basictep to better predict climate change impacts on forest ecosystemsBeniston 2002)
Elevational gradients have been reported as strategic sites to
ssess climatendashtree growth relationships because temperature andrecipitation vary along with elevation gradients and so do veg-tation responses Climate change has resulted in a significant
lowast Corresponding author at Facultad de Ciencias Forestales Universidad Juaacuterezel Estado de Durango Av Papaloapan y Blvd Durango Durango 34120 Mexicoel +52 618 1301096 fax +52 618 1301148
125-7865$ ndash see front matter copy 2013 Elsevier GmbH All rights reservedttpdxdoiorg101016jdendro201305003
copy 2013 Elsevier GmbH All rights reserved
pward shift in speciesrsquo optimum elevation averaging 29 m perecade in western Europe especially for trees in mountain habi-ats (Lenoir et al 2008) The prediction of forest dynamics in theontext of future climate is facilitated by an understanding of treerowth changes related to altitudinal gradients (Zhang and Hebda004) Dendrochronology provides tools to establish a relation-hip between tree growth and dynamic climate variables over timeFritts 2001)
In forests of Durango Mexico dendrochronological analysis haseen used to explore variability in ecosystem response to climateleaveland et al (2003) pointed out that inter-annual rainfall is
argely controlled by large scale atmospheric circulation patternsie El Nino Southern Oscillation) Gonzaacutelez-Elizondo et al (2005)eveloped a comparative study of climatendashgrowth relationships
n high altitudes Villanueva et al (2009) carried out a paleocli-atic study to determine historical hydroclimate variability All
hese studies were developed with Pseudotsuga menziesii since its highly sensitive to climate in the study area However to ournowledge there are no studies on the growth response of Pinusooperi and its relationship to climate at different elevations
P cooperi is a widely distributed species in the Sierra Madreccidental from the southern part of Chihuahua to the center of
urango from 2000 to 2800 m in elevation (Gonzaacutelez-Elizondot al 2007) This species also provides the highest volume andalue to the timber industry in the State of Durango because ofts abundance high wood quality and clean long boles (Cruz et al
rochr
2m(fitg
cpgagh
M
D
taePAcsasLiT
D
dcwoclh
pgo
dfislufat1rtb
C
f(rMtwmpmvTtTttaS
ascs(grf
TD
M Pompa-Garciacutea et al Dend
008) However it is unknown how this species grows in differenticroclimates which would help us predict different responses
extinction migration or adjustment of physiological processes) touture environmental conditions (Viveros-Viveros et al 2009) Its necessary to understand whether there are specific climatic fac-ors that regulate growth variation of P cooperi populations acrossradients
To investigate these issues a dendroclimatological study wasarried out to quantify the relationships between radial growthrecipitation and temperature for several sites across an altitudinalradient We hypothesized that the response to the climate vari-bles would vary systematically along the elevational range withrowth increasing with higher precipitation and decreasing withigher temperature
aterials and methods
escription of the study area
We selected a study region located in the Sierra Madre Occiden-al in northwest Durango Mexico Ecosystems of the study regionre biogeographically important and biologically diverse The for-st composition was mixed with a dominant tree layer of P cooperiinus duranguensis Pinus arizonica Pinus ayacahuite P menziesiibies durangensis Arbutus xalepensis and several species of Quer-us spp (Gonzaacutelez-Elizondo et al 2007) The climate is temperateemi-cool with a long cool summer sub-humid the soils typesre cambisols lithosols regosols and feozems (Pompa 2012) Thetudy area was stratified in five elevational categories Lowest (LW)ow (L) Mid (M) High (H) and Highest (HE) characterized by sim-lar stand structure ecological conditions and forest managementhe sample plot elevation ranged from 2330 to 2905 m
ata sampling and chronology construction
We selected five representative stands along an elevational gra-ient where there were healthy dominant P cooperi trees in theanopy We avoided injured or deformed trees as well as thosehose growth may have been influenced by competition for light
r soil nutrients (Table 1) This strategy was adopted in order toharacterize the climatic response of the majority of the tree popu-ations within the study zones and not just trees that are underigh environmental stress (Wilmking et al 2004)
The field work was carried out in October 2011 Tree ring sam-les were collected from 8 to 16 trees per site along the altitudinalradient At least two increment cores were extracted for each treef P cooperi at 13 m height using increment borers
smso
able 1escriptive data of sampled plots of Pinus cooperi
Site Extreme geographic coordinates Mean
Longitude W () Latitude N ()
HE 105535421 23748323 2905
105534172 23747250
H 105534743 23751968 2813
105533452 23750428
M 105504792 23742671 2680
105502877 23741591
L 105465580 24191390 2424
105377620 24177010
LW 105056047 24141570 2330
105021081 24078309
onologia 31 (2013) 198ndash 204 199
Each of the samples was crossdated using standard den-rochronological techniques to ensure the date of the exact year oformation of each growth ring (Stokes and Smiley 1968) The qual-ty of the dating and measurement of each growth ring was verifiedtatistically using the program COFECHA (Holmes 1983) The bio-ogical and geometric trends not related to climate were removedsing ARSTAN (Cook and Holmes 1984) A cubic smoothing splineunction was fitted to the ring-width series Then dividing eachnnual value of measurement by the value obtained from the curvehe residual index series with a mean of 10 and variance of 0 (Cook987) was generated The result was a residual tree ring index (TRI)epresenting the common signal for the site without autocorrela-ion (Fritts 2001) The TRI was then used to analyze relationshipsetween growth and selected climate variables
limate data and statistical analysis
In order to study of tree growth with climate variables recordsrom 1946 to 2010 were derived from ldquoEl Saltordquo weather stationCNA 2012) because of its proximity to the sample plots (distancesanging from 26 for closer study sites and 46 km for farther ones)ost rainfall occurred during the summer period when 76 of
he total annual precipitation was accumulated and the rest (24)as recorded during the winter (Fig 1) Climatic variables wereonthly total precipitation (P) in mm maximum monthly tem-
erature (Tmax) mean monthly temperature (Tmean) and minimumonthly temperature (Tmin) in C Twelve additional composite
ariables were also included in the analysis previous summer Pmax Tmean and Tmin (PSP PSTmax PSTmean PSTmin respectively) fromhe previous June to the previous September previous winter Pmax Tmean and Tmin (PWP PWTmax PWTmean and PWTmin respec-ively) from the previous October to the current February andhe current summer P Tmax Tmean and Tmin (CSP CSTmax CSTmean
nd CSTmin respectively) from the current June to the currenteptember (growing season)
Residual tree-ring chronologies and climate relationships werenalyzed with DendroClim2002 (Biondi and Waikul 2004) thisoftware computes the statistical significance of the correlationoefficients by calculating 95 quantile limits based on 1000 boot-trapped resamples of the data The statistical software SASSTATreg
SAS 2004) was used to compute the response functions of treerowth to climate by means of a multiple regression A stepwiseegression was then carried out with a level of significance of 010or those climatic variables significantly correlated with TRI The
tepwise method looks at all the variables already included in theodel and deletes any variable that does not produce an F statistic
ignificant at the required level (SAS 2004) To reduce problemsf collinearity among variables in the stepwise regression analysis
elevation (masl) Number of trees Mean Dbh (cm)
10 472
10 501
8 436
16 4625
10 482
200 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
F (1946T respec
(wlm
R
T
fif
fTwsVT
g
TD
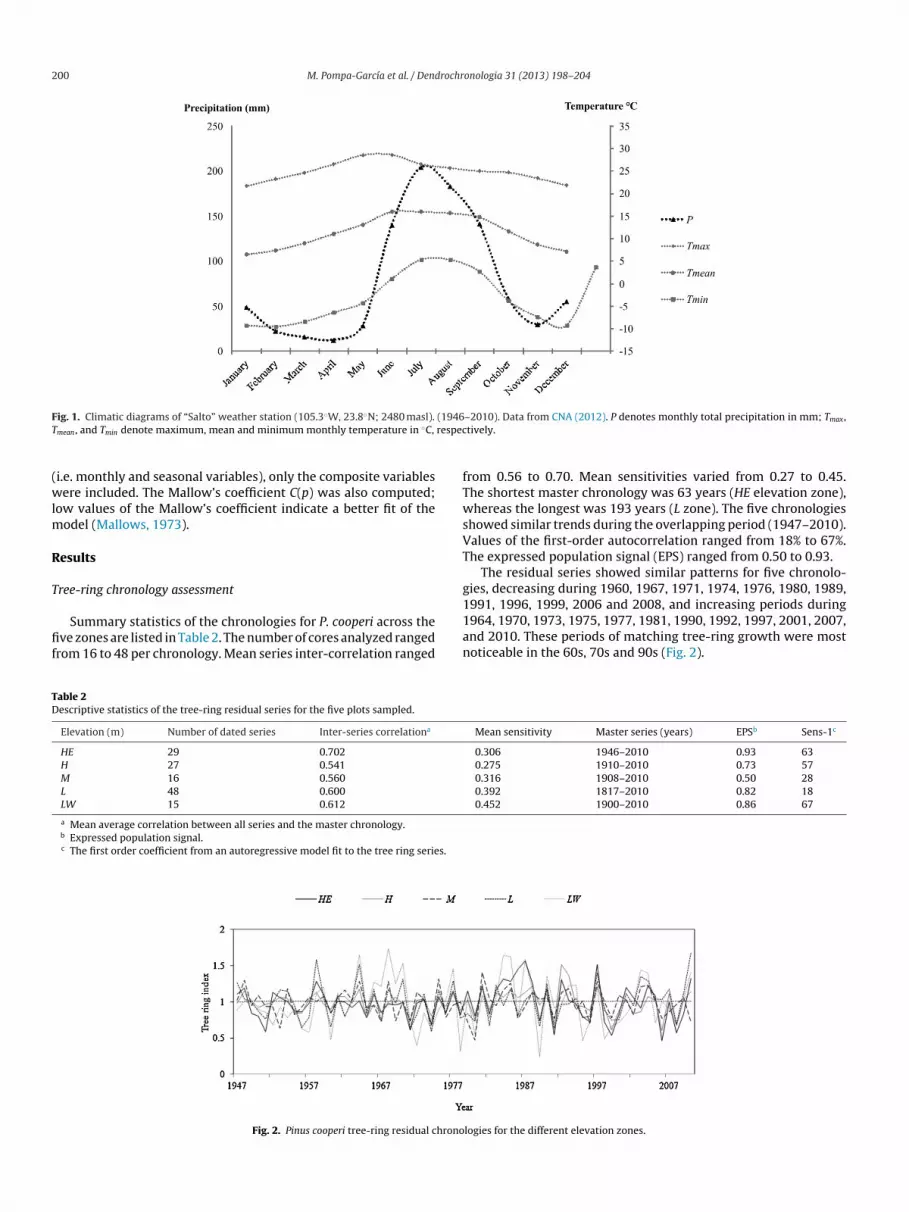
ig 1 Climatic diagrams of ldquoSaltordquo weather station (1053W 238N 2480 masl)
mean and Tmin denote maximum mean and minimum monthly temperature in C
ie monthly and seasonal variables) only the composite variablesere included The Mallowrsquos coefficient C(p) was also computed
ow values of the Mallowrsquos coefficient indicate a better fit of theodel (Mallows 1973)
esults
ree-ring chronology assessment
Summary statistics of the chronologies for P cooperi across theve zones are listed in Table 2 The number of cores analyzed ranged
rom 16 to 48 per chronology Mean series inter-correlation ranged
11an
able 2escriptive statistics of the tree-ring residual series for the five plots sampled
Elevation (m) Number of dated series Inter-series correlationa
HE 29 0702
H 27 0541
M 16 0560
L 48 0600
LW 15 0612
a Mean average correlation between all series and the master chronologyb Expressed population signalc The first order coefficient from an autoregressive model fit to the tree ring series
Fig 2 Pinus cooperi tree-ring residual chrono
ndash2010) Data from CNA (2012) P denotes monthly total precipitation in mm Tmax tively
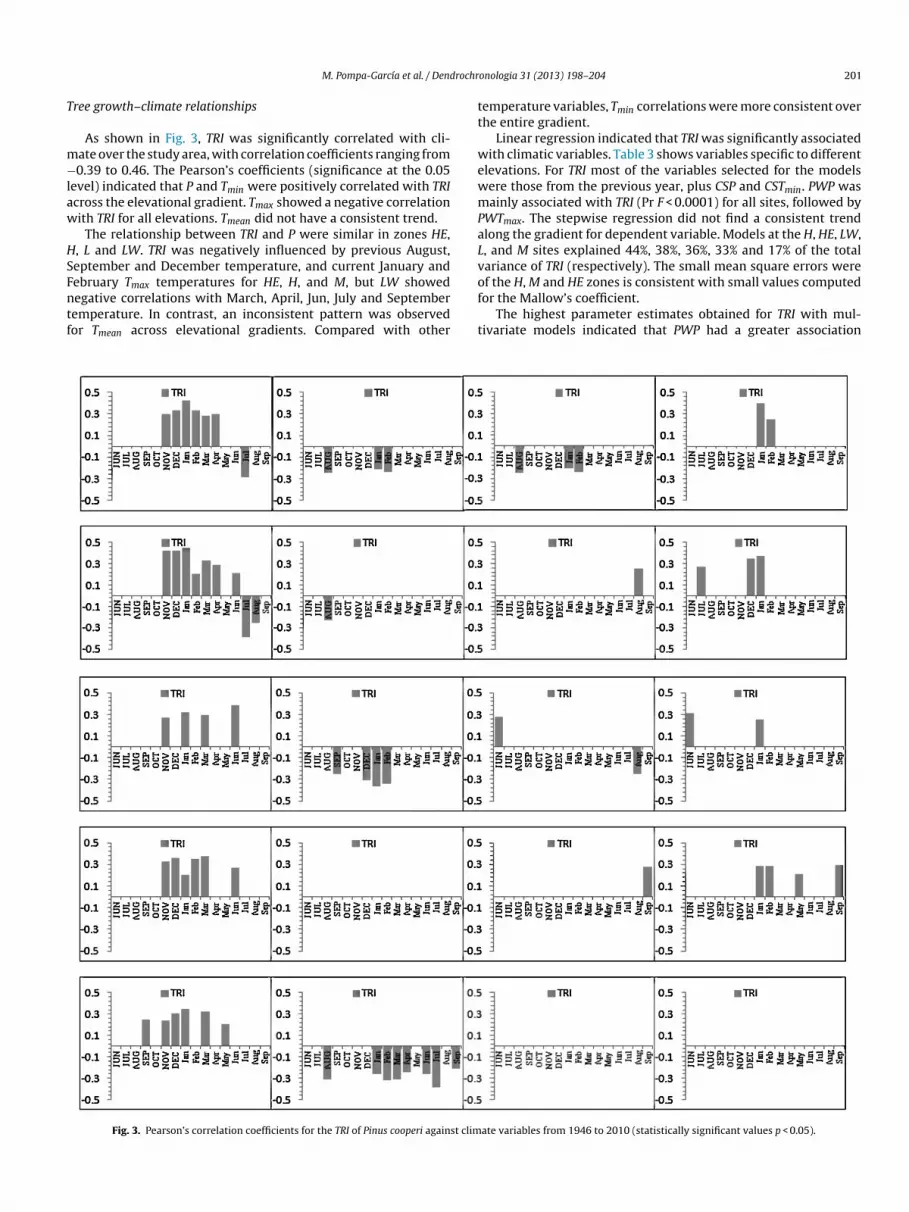
rom 056 to 070 Mean sensitivities varied from 027 to 045he shortest master chronology was 63 years (HE elevation zone)hereas the longest was 193 years (L zone) The five chronologies
howed similar trends during the overlapping period (1947ndash2010)alues of the first-order autocorrelation ranged from 18 to 67he expressed population signal (EPS) ranged from 050 to 093
The residual series showed similar patterns for five chronolo-ies decreasing during 1960 1967 1971 1974 1976 1980 1989
991 1996 1999 2006 and 2008 and increasing periods during964 1970 1973 1975 1977 1981 1990 1992 1997 2001 2007nd 2010 These periods of matching tree-ring growth were mostoticeable in the 60s 70s and 90s (Fig 2)
Mean sensitivity Master series (years) EPSb Sens-1c
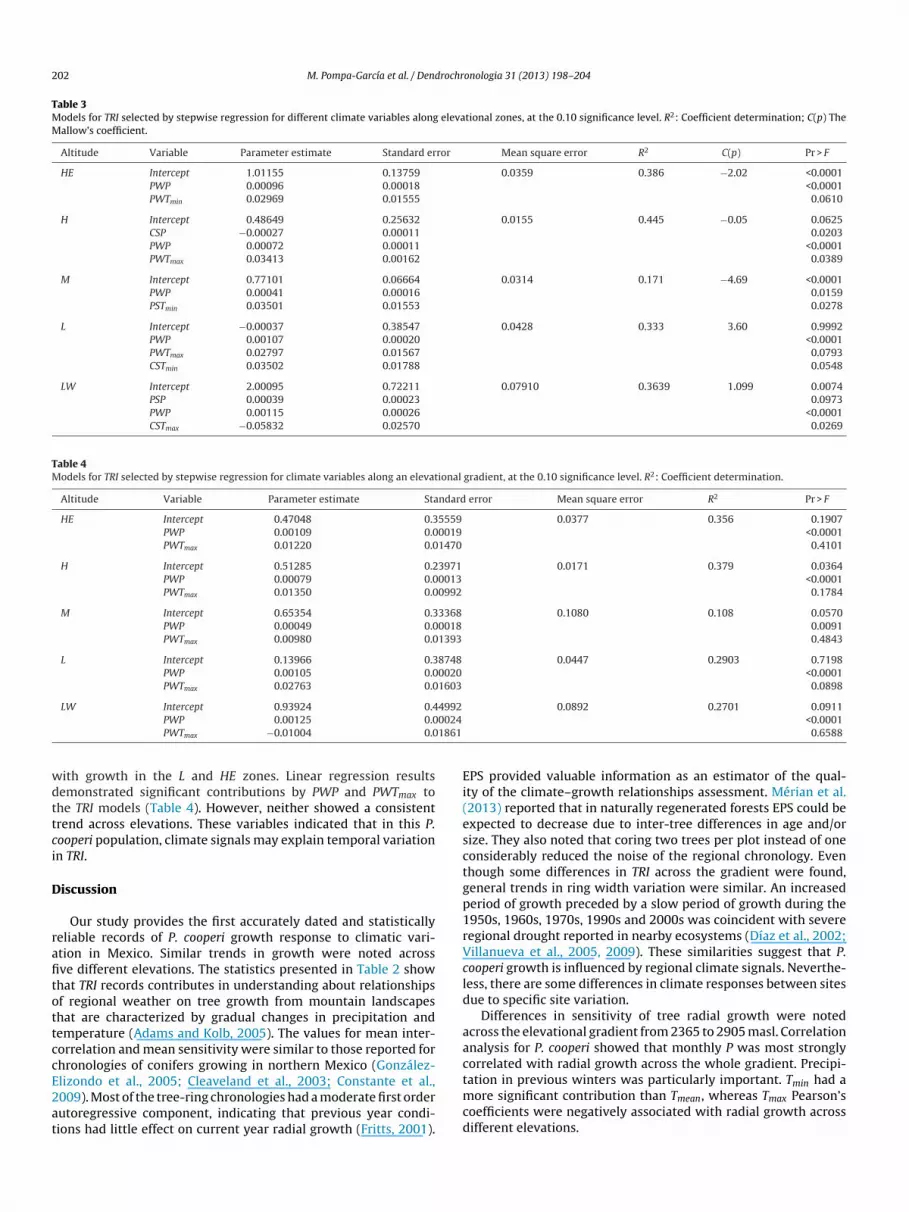
As shown in Fig 3 TRI was significantly correlated with cli-ate over the study area with correlation coefficients ranging from039 to 046 The Pearsonrsquos coefficients (significance at the 005
evel) indicated that P and Tmin were positively correlated with TRIcross the elevational gradient Tmax showed a negative correlationith TRI for all elevations Tmean did not have a consistent trend
The relationship between TRI and P were similar in zones HE L and LW TRI was negatively influenced by previous Augusteptember and December temperature and current January andebruary Tmax temperatures for HE H and M but LW showed
egative correlations with March April Jun July and Septemberemperature In contrast an inconsistent pattern was observedor Tmean across elevational gradients Compared with other
f
t
Fig 3 Pearsonrsquos correlation coefficients for the TRI of Pinus cooperi against clim
onologia 31 (2013) 198ndash 204 201
emperature variables Tmin correlations were more consistent overhe entire gradient
Linear regression indicated that TRI was significantly associatedith climatic variables Table 3 shows variables specific to different
levations For TRI most of the variables selected for the modelsere those from the previous year plus CSP and CSTmin PWP wasainly associated with TRI (Pr F lt 00001) for all sites followed by
WTmax The stepwise regression did not find a consistent trendlong the gradient for dependent variable Models at the H HE LW and M sites explained 44 38 36 33 and 17 of the totalariance of TRI (respectively) The small mean square errors weref the H M and HE zones is consistent with small values computed
or the Mallowrsquos coefficient
The highest parameter estimates obtained for TRI with mul-ivariate models indicated that PWP had a greater association
ate variables from 1946 to 2010 (statistically significant values p lt 005)
202 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
Table 3Models for TRI selected by stepwise regression for different climate variables along elevational zones at the 010 significance level R2 Coefficient determination C(p) TheMallowrsquos coefficient
Altitude Variable Parameter estimate Standard error Mean square error R2 C(p) Pr gt F
Table 4Models for TRI selected by stepwise regression for climate variables along an elevational gradient at the 010 significance level R2 Coefficient determination
Altitude Variable Parameter estimate Standard error Mean square error R2 Pr gt F
ith growth in the L and HE zones Linear regression resultsemonstrated significant contributions by PWP and PWTmax tohe TRI models (Table 4) However neither showed a consistentrend across elevations These variables indicated that in this Pooperi population climate signals may explain temporal variationn TRI
iscussion
Our study provides the first accurately dated and statisticallyeliable records of P cooperi growth response to climatic vari-tion in Mexico Similar trends in growth were noted acrossve different elevations The statistics presented in Table 2 showhat TRI records contributes in understanding about relationshipsf regional weather on tree growth from mountain landscapeshat are characterized by gradual changes in precipitation andemperature (Adams and Kolb 2005) The values for mean inter-orrelation and mean sensitivity were similar to those reported forhronologies of conifers growing in northern Mexico (Gonzaacutelez-
lizondo et al 2005 Cleaveland et al 2003 Constante et al009) Most of the tree-ring chronologies had a moderate first orderutoregressive component indicating that previous year condi-ions had little effect on current year radial growth (Fritts 2001)
tmcd
lt0000106588
PS provided valuable information as an estimator of the qual-ty of the climatendashgrowth relationships assessment Meacuterian et al2013) reported that in naturally regenerated forests EPS could bexpected to decrease due to inter-tree differences in age andorize They also noted that coring two trees per plot instead of oneonsiderably reduced the noise of the regional chronology Evenhough some differences in TRI across the gradient were foundeneral trends in ring width variation were similar An increasederiod of growth preceded by a slow period of growth during the950s 1960s 1970s 1990s and 2000s was coincident with severeegional drought reported in nearby ecosystems (Diacuteaz et al 2002illanueva et al 2005 2009) These similarities suggest that Pooperi growth is influenced by regional climate signals Neverthe-ess there are some differences in climate responses between sitesue to specific site variation
Differences in sensitivity of tree radial growth were notedcross the elevational gradient from 2365 to 2905 masl Correlationnalysis for P cooperi showed that monthly P was most stronglyorrelated with radial growth across the whole gradient Precipi-
ation in previous winters was particularly important Tmin had a
ore significant contribution than Tmean whereas Tmax Pearsonrsquosoefficients were negatively associated with radial growth acrossifferent elevations
rochr
etswsrl(weihf
ettRcl
pLslcdta(
ft(rt
gnMfst(
gptptTbcea(t
wngzi2
sgtomsaagelcghgahse2df2ag
C
aacdgl
cwdggraogtiw
A
MtdCss
M Pompa-Garciacutea et al Dend
The radial growth of P cooperi at all elevations was mainly influ-nced by PWP Tables 3 and 4 suggest that water storage during theree dormancy period is useful as a reserve for the next growing sea-on (Litton et al 2007) This finding supports the hypothesis thatet winters contributed to subsequent warm-season photosynthe-
is and therefore to tree growth (Kerhoulas et al 2013) Winterains are usually of low intensity favoring high infiltration andow evapotranspiration which result in a positive water balanceConstante et al 2009) Also slow winter precipitation rechargesater storage in deep soil horizons where it is protected from soil
vaporation The PWP increases the water stored in soil until a crit-cal level is reached that is required to break bud dormancy andence start tree growth This result is consistent with other results
rom nearby areas (Gonzaacutelez-Elizondo et al 2005)In regards to the negative contribution of CSP to growth at high
levations it could be explained by tree water stress In summerhese sites reach maximum soil water holding capacity affectingree growth due to the activity of soil microorganisms (Bartelt-yser et al 2005) Furthermore Li et al (2012) reported thatloudiness is an important factor that reduces growth through lightimitation
Tmax had a negative influence on TRI suggesting that high tem-eratures may lead to physiological impairment According toebourgeois et al (2012) the optimal photosynthesis rate corre-ponds to medium temperatures An acceleration of warming couldead to reduced tree vigor and increased risk of mortality or sus-eptibility to insect attack Tmax may indicate high vapor pressureeficit which cause the stomata to close thus reducing photosyn-hesis Allocation of carbohydrates is of major importance for leafnd root expansion which affects subsequent radial growth of treesMisson et al 2004)
These findings suggest that yearly growth in the high-elevationorest was limited more by maximum temperatures than meanemperatures This is consistent with findings from Adams and Kolb2005) whose research suggested that an increasing respirationate (ie carbon allocation to respiration) during periods of highemperatures could limit radial growth in temperate ecosystems
There was a strong positive correlation between Tmin and radialrowth There are a number of possible ecophysiological mecha-isms whereby a cold winter could be associated with tree growthinimal temperatures during winter will usually result in soil
reezing and a delay in growth until the ground has thawed in thepring (Chhin et al 2008) The positive response to Tmin is consis-ent with previous studies on other conifers in mountainous areasHuang et al 2010)
Along the whole elevational gradient correlation analysis sug-ested that warmer Tmin enhanced growth Tmin paired with PWPlays a major role as a predictor variable of tree growth par-icularly in higher-elevation stands where it explains a largeercentage of the variance in regression models These interac-ions contribute to prolonging the growing season of P cooperihe underlying mechanism of this effect could be interpreted toe that growth is enhanced by a low evapotranspiration rate inombination with water availability in the upper zones (Rossit al 2008) Together these environmental conditions promoten early budburst cambial reactivation and thus radial growthLebourgeois et al 2012) Lenoir et al (2008) has reported a similarrend
Tree growth at H LW L and HE zones was strongly associatedith climate whereas M zone had less-sensitive chronologies witho clear common response pattern Lower correlation between
rowth and climate variables particularly for lower and drierones has often been interpreted as a strategy to avoid ecophys-ological impairment during the driest period (Lebourgeois et al012)
KaTt
onologia 31 (2013) 198ndash 204 203
The elevational differences in the R2 and parameter regres-ion values did not reflect noteworthy elevational trends in radialrowth Indeed there were no discernible differences across eleva-ions in the radial growth pattern or climate growth relationshipsf P cooperi This is inconsistent with the concept of differing cli-atic signals at different elevations captured in tree ring data as
hown by Bickford et al (2011) Instead the differences we foundre probably due to specific site conditions The lack of system-tic trends across the elevational gradient is also due to the uniqueeography ecology and topography of the sites sampled (Tardift al 2003) in contrast with the ideal area for dendroclimato-ogical studies reported by Li et al (2012) Another explanationould be that trees responded to the climatic signals based onenetic adaptations throughout their range (Prior et al 2012) Itas been demonstrated that coniferous populations differentiateenetically in response to differential selection pressure occurringlong the elevational gradient The populations of lower altitudesave lower cold tolerance and higher growth potential (earlierhoot elongation later growth cessation and larger total shootlongation) than those of higher altitudes (Viveros-Viveros et al009) Dendrochronology approaches may have the potential toetect differences among populations to detect suitable genotypesor forest management strategies under climate change (Chen et al010) Stan and Daniels (2010) found that the complementarypplication of basal area improves the predictive capacity of treerowth rings in coastal British Columbia
onclusion
The results in this study did not indicate systematic trends alongn elevational gradient in tree growth response to climatic vari-tion in contrast with our hypothesis However some commonlimatic variables independent of local microclimate factors wererivers in regression models across the elevational zones Perhapsenetic adaptations have occurred as a response of populations toocal environmental variation
Radial tree growth was mainly correlated with previous climateonditions Tree growth was positively and significantly correlatedith previous winter conditions at most of the sites along the gra-ient Wet and cold winters were positively associated with radialrowth at most elevations whereas warm winters affected treerowth negatively The results showed that tree growth did notespond to precipitation or temperature as independent variables Pnd T were useful explanatory variables indicating the importancef future climate change which could significantly influence treerowth during the 21st century These projections have implica-ions for P cooperi radial growth For example a potential reductionn radial growth could be expected in response to less abundant
inter precipitation plus higher temperatures
cknowledgments
This work was conducted within the framework of Academicobility supported by the UJED amp CONACyT ldquoEstancias Posdoc-
orales y Sabaacuteticas al Extranjero para la Consolidacioacuten de Grupose InvestigacioacutenrdquoldquoPostdoctoral and Sabbatical Stays Abroad foronsolidation of Researchers Teamsrdquo We would like to thank theupport given by School of Forestry at Northern Arizona Univer-ity Also we recognize Larissa Yocom Amanda Stan Thomas E
olb and Jim Allen for their helpful assistance Omar Duraacuten Guerrand Joseacute Santiago Saacutenchez provided help with field data gatheringhe authors are grateful to Editors and anonymous reviewers forheir useful comments and suggestions
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)
rochr
2m(fitg
cpgagh
M
D
taePAcsasLiT
D
dcwoclh
pgo
dfislufat1rtb
C
f(rMtwmpmvTtTttaS
ascs(grf
TD
M Pompa-Garciacutea et al Dend
008) However it is unknown how this species grows in differenticroclimates which would help us predict different responses
extinction migration or adjustment of physiological processes) touture environmental conditions (Viveros-Viveros et al 2009) Its necessary to understand whether there are specific climatic fac-ors that regulate growth variation of P cooperi populations acrossradients
To investigate these issues a dendroclimatological study wasarried out to quantify the relationships between radial growthrecipitation and temperature for several sites across an altitudinalradient We hypothesized that the response to the climate vari-bles would vary systematically along the elevational range withrowth increasing with higher precipitation and decreasing withigher temperature
aterials and methods
escription of the study area
We selected a study region located in the Sierra Madre Occiden-al in northwest Durango Mexico Ecosystems of the study regionre biogeographically important and biologically diverse The for-st composition was mixed with a dominant tree layer of P cooperiinus duranguensis Pinus arizonica Pinus ayacahuite P menziesiibies durangensis Arbutus xalepensis and several species of Quer-us spp (Gonzaacutelez-Elizondo et al 2007) The climate is temperateemi-cool with a long cool summer sub-humid the soils typesre cambisols lithosols regosols and feozems (Pompa 2012) Thetudy area was stratified in five elevational categories Lowest (LW)ow (L) Mid (M) High (H) and Highest (HE) characterized by sim-lar stand structure ecological conditions and forest managementhe sample plot elevation ranged from 2330 to 2905 m
ata sampling and chronology construction
We selected five representative stands along an elevational gra-ient where there were healthy dominant P cooperi trees in theanopy We avoided injured or deformed trees as well as thosehose growth may have been influenced by competition for light
r soil nutrients (Table 1) This strategy was adopted in order toharacterize the climatic response of the majority of the tree popu-ations within the study zones and not just trees that are underigh environmental stress (Wilmking et al 2004)
The field work was carried out in October 2011 Tree ring sam-les were collected from 8 to 16 trees per site along the altitudinalradient At least two increment cores were extracted for each treef P cooperi at 13 m height using increment borers
smso
able 1escriptive data of sampled plots of Pinus cooperi
Site Extreme geographic coordinates Mean
Longitude W () Latitude N ()
HE 105535421 23748323 2905
105534172 23747250
H 105534743 23751968 2813
105533452 23750428
M 105504792 23742671 2680
105502877 23741591
L 105465580 24191390 2424
105377620 24177010
LW 105056047 24141570 2330
105021081 24078309
onologia 31 (2013) 198ndash 204 199
Each of the samples was crossdated using standard den-rochronological techniques to ensure the date of the exact year oformation of each growth ring (Stokes and Smiley 1968) The qual-ty of the dating and measurement of each growth ring was verifiedtatistically using the program COFECHA (Holmes 1983) The bio-ogical and geometric trends not related to climate were removedsing ARSTAN (Cook and Holmes 1984) A cubic smoothing splineunction was fitted to the ring-width series Then dividing eachnnual value of measurement by the value obtained from the curvehe residual index series with a mean of 10 and variance of 0 (Cook987) was generated The result was a residual tree ring index (TRI)epresenting the common signal for the site without autocorrela-ion (Fritts 2001) The TRI was then used to analyze relationshipsetween growth and selected climate variables
limate data and statistical analysis
In order to study of tree growth with climate variables recordsrom 1946 to 2010 were derived from ldquoEl Saltordquo weather stationCNA 2012) because of its proximity to the sample plots (distancesanging from 26 for closer study sites and 46 km for farther ones)ost rainfall occurred during the summer period when 76 of
he total annual precipitation was accumulated and the rest (24)as recorded during the winter (Fig 1) Climatic variables wereonthly total precipitation (P) in mm maximum monthly tem-
erature (Tmax) mean monthly temperature (Tmean) and minimumonthly temperature (Tmin) in C Twelve additional composite
ariables were also included in the analysis previous summer Pmax Tmean and Tmin (PSP PSTmax PSTmean PSTmin respectively) fromhe previous June to the previous September previous winter Pmax Tmean and Tmin (PWP PWTmax PWTmean and PWTmin respec-ively) from the previous October to the current February andhe current summer P Tmax Tmean and Tmin (CSP CSTmax CSTmean
nd CSTmin respectively) from the current June to the currenteptember (growing season)
Residual tree-ring chronologies and climate relationships werenalyzed with DendroClim2002 (Biondi and Waikul 2004) thisoftware computes the statistical significance of the correlationoefficients by calculating 95 quantile limits based on 1000 boot-trapped resamples of the data The statistical software SASSTATreg
SAS 2004) was used to compute the response functions of treerowth to climate by means of a multiple regression A stepwiseegression was then carried out with a level of significance of 010or those climatic variables significantly correlated with TRI The
tepwise method looks at all the variables already included in theodel and deletes any variable that does not produce an F statistic
ignificant at the required level (SAS 2004) To reduce problemsf collinearity among variables in the stepwise regression analysis
elevation (masl) Number of trees Mean Dbh (cm)
10 472
10 501
8 436
16 4625
10 482
200 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
F (1946T respec
(wlm
R
T
fif
fTwsVT
g
TD
ig 1 Climatic diagrams of ldquoSaltordquo weather station (1053W 238N 2480 masl)
mean and Tmin denote maximum mean and minimum monthly temperature in C
ie monthly and seasonal variables) only the composite variablesere included The Mallowrsquos coefficient C(p) was also computed
ow values of the Mallowrsquos coefficient indicate a better fit of theodel (Mallows 1973)
esults
ree-ring chronology assessment
Summary statistics of the chronologies for P cooperi across theve zones are listed in Table 2 The number of cores analyzed ranged
rom 16 to 48 per chronology Mean series inter-correlation ranged
11an
able 2escriptive statistics of the tree-ring residual series for the five plots sampled
Elevation (m) Number of dated series Inter-series correlationa
HE 29 0702
H 27 0541
M 16 0560
L 48 0600
LW 15 0612
a Mean average correlation between all series and the master chronologyb Expressed population signalc The first order coefficient from an autoregressive model fit to the tree ring series
Fig 2 Pinus cooperi tree-ring residual chrono
ndash2010) Data from CNA (2012) P denotes monthly total precipitation in mm Tmax tively
rom 056 to 070 Mean sensitivities varied from 027 to 045he shortest master chronology was 63 years (HE elevation zone)hereas the longest was 193 years (L zone) The five chronologies
howed similar trends during the overlapping period (1947ndash2010)alues of the first-order autocorrelation ranged from 18 to 67he expressed population signal (EPS) ranged from 050 to 093
The residual series showed similar patterns for five chronolo-ies decreasing during 1960 1967 1971 1974 1976 1980 1989
991 1996 1999 2006 and 2008 and increasing periods during964 1970 1973 1975 1977 1981 1990 1992 1997 2001 2007nd 2010 These periods of matching tree-ring growth were mostoticeable in the 60s 70s and 90s (Fig 2)
Mean sensitivity Master series (years) EPSb Sens-1c
As shown in Fig 3 TRI was significantly correlated with cli-ate over the study area with correlation coefficients ranging from039 to 046 The Pearsonrsquos coefficients (significance at the 005
evel) indicated that P and Tmin were positively correlated with TRIcross the elevational gradient Tmax showed a negative correlationith TRI for all elevations Tmean did not have a consistent trend
The relationship between TRI and P were similar in zones HE L and LW TRI was negatively influenced by previous Augusteptember and December temperature and current January andebruary Tmax temperatures for HE H and M but LW showed
egative correlations with March April Jun July and Septemberemperature In contrast an inconsistent pattern was observedor Tmean across elevational gradients Compared with other
f
t
Fig 3 Pearsonrsquos correlation coefficients for the TRI of Pinus cooperi against clim
onologia 31 (2013) 198ndash 204 201
emperature variables Tmin correlations were more consistent overhe entire gradient
Linear regression indicated that TRI was significantly associatedith climatic variables Table 3 shows variables specific to different
levations For TRI most of the variables selected for the modelsere those from the previous year plus CSP and CSTmin PWP wasainly associated with TRI (Pr F lt 00001) for all sites followed by
WTmax The stepwise regression did not find a consistent trendlong the gradient for dependent variable Models at the H HE LW and M sites explained 44 38 36 33 and 17 of the totalariance of TRI (respectively) The small mean square errors weref the H M and HE zones is consistent with small values computed
or the Mallowrsquos coefficient
The highest parameter estimates obtained for TRI with mul-ivariate models indicated that PWP had a greater association
ate variables from 1946 to 2010 (statistically significant values p lt 005)
202 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
Table 3Models for TRI selected by stepwise regression for different climate variables along elevational zones at the 010 significance level R2 Coefficient determination C(p) TheMallowrsquos coefficient
Altitude Variable Parameter estimate Standard error Mean square error R2 C(p) Pr gt F
Table 4Models for TRI selected by stepwise regression for climate variables along an elevational gradient at the 010 significance level R2 Coefficient determination
Altitude Variable Parameter estimate Standard error Mean square error R2 Pr gt F
ith growth in the L and HE zones Linear regression resultsemonstrated significant contributions by PWP and PWTmax tohe TRI models (Table 4) However neither showed a consistentrend across elevations These variables indicated that in this Pooperi population climate signals may explain temporal variationn TRI
iscussion
Our study provides the first accurately dated and statisticallyeliable records of P cooperi growth response to climatic vari-tion in Mexico Similar trends in growth were noted acrossve different elevations The statistics presented in Table 2 showhat TRI records contributes in understanding about relationshipsf regional weather on tree growth from mountain landscapeshat are characterized by gradual changes in precipitation andemperature (Adams and Kolb 2005) The values for mean inter-orrelation and mean sensitivity were similar to those reported forhronologies of conifers growing in northern Mexico (Gonzaacutelez-
lizondo et al 2005 Cleaveland et al 2003 Constante et al009) Most of the tree-ring chronologies had a moderate first orderutoregressive component indicating that previous year condi-ions had little effect on current year radial growth (Fritts 2001)
tmcd
lt0000106588
PS provided valuable information as an estimator of the qual-ty of the climatendashgrowth relationships assessment Meacuterian et al2013) reported that in naturally regenerated forests EPS could bexpected to decrease due to inter-tree differences in age andorize They also noted that coring two trees per plot instead of oneonsiderably reduced the noise of the regional chronology Evenhough some differences in TRI across the gradient were foundeneral trends in ring width variation were similar An increasederiod of growth preceded by a slow period of growth during the950s 1960s 1970s 1990s and 2000s was coincident with severeegional drought reported in nearby ecosystems (Diacuteaz et al 2002illanueva et al 2005 2009) These similarities suggest that Pooperi growth is influenced by regional climate signals Neverthe-ess there are some differences in climate responses between sitesue to specific site variation
Differences in sensitivity of tree radial growth were notedcross the elevational gradient from 2365 to 2905 masl Correlationnalysis for P cooperi showed that monthly P was most stronglyorrelated with radial growth across the whole gradient Precipi-
ation in previous winters was particularly important Tmin had a
ore significant contribution than Tmean whereas Tmax Pearsonrsquosoefficients were negatively associated with radial growth acrossifferent elevations
rochr
etswsrl(weihf
ettRcl
pLslcdta(
ft(rt
gnMfst(
gptptTbcea(t
wngzi2
sgtomsaagelcghgahse2df2ag
C
aacdgl
cwdggraogtiw
A
MtdCss
M Pompa-Garciacutea et al Dend
The radial growth of P cooperi at all elevations was mainly influ-nced by PWP Tables 3 and 4 suggest that water storage during theree dormancy period is useful as a reserve for the next growing sea-on (Litton et al 2007) This finding supports the hypothesis thatet winters contributed to subsequent warm-season photosynthe-
is and therefore to tree growth (Kerhoulas et al 2013) Winterains are usually of low intensity favoring high infiltration andow evapotranspiration which result in a positive water balanceConstante et al 2009) Also slow winter precipitation rechargesater storage in deep soil horizons where it is protected from soil
vaporation The PWP increases the water stored in soil until a crit-cal level is reached that is required to break bud dormancy andence start tree growth This result is consistent with other results
rom nearby areas (Gonzaacutelez-Elizondo et al 2005)In regards to the negative contribution of CSP to growth at high
levations it could be explained by tree water stress In summerhese sites reach maximum soil water holding capacity affectingree growth due to the activity of soil microorganisms (Bartelt-yser et al 2005) Furthermore Li et al (2012) reported thatloudiness is an important factor that reduces growth through lightimitation
Tmax had a negative influence on TRI suggesting that high tem-eratures may lead to physiological impairment According toebourgeois et al (2012) the optimal photosynthesis rate corre-ponds to medium temperatures An acceleration of warming couldead to reduced tree vigor and increased risk of mortality or sus-eptibility to insect attack Tmax may indicate high vapor pressureeficit which cause the stomata to close thus reducing photosyn-hesis Allocation of carbohydrates is of major importance for leafnd root expansion which affects subsequent radial growth of treesMisson et al 2004)
These findings suggest that yearly growth in the high-elevationorest was limited more by maximum temperatures than meanemperatures This is consistent with findings from Adams and Kolb2005) whose research suggested that an increasing respirationate (ie carbon allocation to respiration) during periods of highemperatures could limit radial growth in temperate ecosystems
There was a strong positive correlation between Tmin and radialrowth There are a number of possible ecophysiological mecha-isms whereby a cold winter could be associated with tree growthinimal temperatures during winter will usually result in soil
reezing and a delay in growth until the ground has thawed in thepring (Chhin et al 2008) The positive response to Tmin is consis-ent with previous studies on other conifers in mountainous areasHuang et al 2010)
Along the whole elevational gradient correlation analysis sug-ested that warmer Tmin enhanced growth Tmin paired with PWPlays a major role as a predictor variable of tree growth par-icularly in higher-elevation stands where it explains a largeercentage of the variance in regression models These interac-ions contribute to prolonging the growing season of P cooperihe underlying mechanism of this effect could be interpreted toe that growth is enhanced by a low evapotranspiration rate inombination with water availability in the upper zones (Rossit al 2008) Together these environmental conditions promoten early budburst cambial reactivation and thus radial growthLebourgeois et al 2012) Lenoir et al (2008) has reported a similarrend
Tree growth at H LW L and HE zones was strongly associatedith climate whereas M zone had less-sensitive chronologies witho clear common response pattern Lower correlation between
rowth and climate variables particularly for lower and drierones has often been interpreted as a strategy to avoid ecophys-ological impairment during the driest period (Lebourgeois et al012)
KaTt
onologia 31 (2013) 198ndash 204 203
The elevational differences in the R2 and parameter regres-ion values did not reflect noteworthy elevational trends in radialrowth Indeed there were no discernible differences across eleva-ions in the radial growth pattern or climate growth relationshipsf P cooperi This is inconsistent with the concept of differing cli-atic signals at different elevations captured in tree ring data as
hown by Bickford et al (2011) Instead the differences we foundre probably due to specific site conditions The lack of system-tic trends across the elevational gradient is also due to the uniqueeography ecology and topography of the sites sampled (Tardift al 2003) in contrast with the ideal area for dendroclimato-ogical studies reported by Li et al (2012) Another explanationould be that trees responded to the climatic signals based onenetic adaptations throughout their range (Prior et al 2012) Itas been demonstrated that coniferous populations differentiateenetically in response to differential selection pressure occurringlong the elevational gradient The populations of lower altitudesave lower cold tolerance and higher growth potential (earlierhoot elongation later growth cessation and larger total shootlongation) than those of higher altitudes (Viveros-Viveros et al009) Dendrochronology approaches may have the potential toetect differences among populations to detect suitable genotypesor forest management strategies under climate change (Chen et al010) Stan and Daniels (2010) found that the complementarypplication of basal area improves the predictive capacity of treerowth rings in coastal British Columbia
onclusion
The results in this study did not indicate systematic trends alongn elevational gradient in tree growth response to climatic vari-tion in contrast with our hypothesis However some commonlimatic variables independent of local microclimate factors wererivers in regression models across the elevational zones Perhapsenetic adaptations have occurred as a response of populations toocal environmental variation
Radial tree growth was mainly correlated with previous climateonditions Tree growth was positively and significantly correlatedith previous winter conditions at most of the sites along the gra-ient Wet and cold winters were positively associated with radialrowth at most elevations whereas warm winters affected treerowth negatively The results showed that tree growth did notespond to precipitation or temperature as independent variables Pnd T were useful explanatory variables indicating the importancef future climate change which could significantly influence treerowth during the 21st century These projections have implica-ions for P cooperi radial growth For example a potential reductionn radial growth could be expected in response to less abundant
inter precipitation plus higher temperatures
cknowledgments
This work was conducted within the framework of Academicobility supported by the UJED amp CONACyT ldquoEstancias Posdoc-
orales y Sabaacuteticas al Extranjero para la Consolidacioacuten de Grupose InvestigacioacutenrdquoldquoPostdoctoral and Sabbatical Stays Abroad foronsolidation of Researchers Teamsrdquo We would like to thank theupport given by School of Forestry at Northern Arizona Univer-ity Also we recognize Larissa Yocom Amanda Stan Thomas E
olb and Jim Allen for their helpful assistance Omar Duraacuten Guerrand Joseacute Santiago Saacutenchez provided help with field data gatheringhe authors are grateful to Editors and anonymous reviewers forheir useful comments and suggestions
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)
200 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
F (1946T respec
(wlm
R
T
fif
fTwsVT
g
TD
ig 1 Climatic diagrams of ldquoSaltordquo weather station (1053W 238N 2480 masl)
mean and Tmin denote maximum mean and minimum monthly temperature in C
ie monthly and seasonal variables) only the composite variablesere included The Mallowrsquos coefficient C(p) was also computed
ow values of the Mallowrsquos coefficient indicate a better fit of theodel (Mallows 1973)
esults
ree-ring chronology assessment
Summary statistics of the chronologies for P cooperi across theve zones are listed in Table 2 The number of cores analyzed ranged
rom 16 to 48 per chronology Mean series inter-correlation ranged
11an
able 2escriptive statistics of the tree-ring residual series for the five plots sampled
Elevation (m) Number of dated series Inter-series correlationa
HE 29 0702
H 27 0541
M 16 0560
L 48 0600
LW 15 0612
a Mean average correlation between all series and the master chronologyb Expressed population signalc The first order coefficient from an autoregressive model fit to the tree ring series
Fig 2 Pinus cooperi tree-ring residual chrono
ndash2010) Data from CNA (2012) P denotes monthly total precipitation in mm Tmax tively
rom 056 to 070 Mean sensitivities varied from 027 to 045he shortest master chronology was 63 years (HE elevation zone)hereas the longest was 193 years (L zone) The five chronologies
howed similar trends during the overlapping period (1947ndash2010)alues of the first-order autocorrelation ranged from 18 to 67he expressed population signal (EPS) ranged from 050 to 093
The residual series showed similar patterns for five chronolo-ies decreasing during 1960 1967 1971 1974 1976 1980 1989
991 1996 1999 2006 and 2008 and increasing periods during964 1970 1973 1975 1977 1981 1990 1992 1997 2001 2007nd 2010 These periods of matching tree-ring growth were mostoticeable in the 60s 70s and 90s (Fig 2)
Mean sensitivity Master series (years) EPSb Sens-1c
As shown in Fig 3 TRI was significantly correlated with cli-ate over the study area with correlation coefficients ranging from039 to 046 The Pearsonrsquos coefficients (significance at the 005
evel) indicated that P and Tmin were positively correlated with TRIcross the elevational gradient Tmax showed a negative correlationith TRI for all elevations Tmean did not have a consistent trend
The relationship between TRI and P were similar in zones HE L and LW TRI was negatively influenced by previous Augusteptember and December temperature and current January andebruary Tmax temperatures for HE H and M but LW showed
egative correlations with March April Jun July and Septemberemperature In contrast an inconsistent pattern was observedor Tmean across elevational gradients Compared with other
f
t
Fig 3 Pearsonrsquos correlation coefficients for the TRI of Pinus cooperi against clim
onologia 31 (2013) 198ndash 204 201
emperature variables Tmin correlations were more consistent overhe entire gradient
Linear regression indicated that TRI was significantly associatedith climatic variables Table 3 shows variables specific to different
levations For TRI most of the variables selected for the modelsere those from the previous year plus CSP and CSTmin PWP wasainly associated with TRI (Pr F lt 00001) for all sites followed by
WTmax The stepwise regression did not find a consistent trendlong the gradient for dependent variable Models at the H HE LW and M sites explained 44 38 36 33 and 17 of the totalariance of TRI (respectively) The small mean square errors weref the H M and HE zones is consistent with small values computed
or the Mallowrsquos coefficient
The highest parameter estimates obtained for TRI with mul-ivariate models indicated that PWP had a greater association
ate variables from 1946 to 2010 (statistically significant values p lt 005)
202 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
Table 3Models for TRI selected by stepwise regression for different climate variables along elevational zones at the 010 significance level R2 Coefficient determination C(p) TheMallowrsquos coefficient
Altitude Variable Parameter estimate Standard error Mean square error R2 C(p) Pr gt F
Table 4Models for TRI selected by stepwise regression for climate variables along an elevational gradient at the 010 significance level R2 Coefficient determination
Altitude Variable Parameter estimate Standard error Mean square error R2 Pr gt F
ith growth in the L and HE zones Linear regression resultsemonstrated significant contributions by PWP and PWTmax tohe TRI models (Table 4) However neither showed a consistentrend across elevations These variables indicated that in this Pooperi population climate signals may explain temporal variationn TRI
iscussion
Our study provides the first accurately dated and statisticallyeliable records of P cooperi growth response to climatic vari-tion in Mexico Similar trends in growth were noted acrossve different elevations The statistics presented in Table 2 showhat TRI records contributes in understanding about relationshipsf regional weather on tree growth from mountain landscapeshat are characterized by gradual changes in precipitation andemperature (Adams and Kolb 2005) The values for mean inter-orrelation and mean sensitivity were similar to those reported forhronologies of conifers growing in northern Mexico (Gonzaacutelez-
lizondo et al 2005 Cleaveland et al 2003 Constante et al009) Most of the tree-ring chronologies had a moderate first orderutoregressive component indicating that previous year condi-ions had little effect on current year radial growth (Fritts 2001)
tmcd
lt0000106588
PS provided valuable information as an estimator of the qual-ty of the climatendashgrowth relationships assessment Meacuterian et al2013) reported that in naturally regenerated forests EPS could bexpected to decrease due to inter-tree differences in age andorize They also noted that coring two trees per plot instead of oneonsiderably reduced the noise of the regional chronology Evenhough some differences in TRI across the gradient were foundeneral trends in ring width variation were similar An increasederiod of growth preceded by a slow period of growth during the950s 1960s 1970s 1990s and 2000s was coincident with severeegional drought reported in nearby ecosystems (Diacuteaz et al 2002illanueva et al 2005 2009) These similarities suggest that Pooperi growth is influenced by regional climate signals Neverthe-ess there are some differences in climate responses between sitesue to specific site variation
Differences in sensitivity of tree radial growth were notedcross the elevational gradient from 2365 to 2905 masl Correlationnalysis for P cooperi showed that monthly P was most stronglyorrelated with radial growth across the whole gradient Precipi-
ation in previous winters was particularly important Tmin had a
ore significant contribution than Tmean whereas Tmax Pearsonrsquosoefficients were negatively associated with radial growth acrossifferent elevations
rochr
etswsrl(weihf
ettRcl
pLslcdta(
ft(rt
gnMfst(
gptptTbcea(t
wngzi2
sgtomsaagelcghgahse2df2ag
C
aacdgl
cwdggraogtiw
A
MtdCss
M Pompa-Garciacutea et al Dend
The radial growth of P cooperi at all elevations was mainly influ-nced by PWP Tables 3 and 4 suggest that water storage during theree dormancy period is useful as a reserve for the next growing sea-on (Litton et al 2007) This finding supports the hypothesis thatet winters contributed to subsequent warm-season photosynthe-
is and therefore to tree growth (Kerhoulas et al 2013) Winterains are usually of low intensity favoring high infiltration andow evapotranspiration which result in a positive water balanceConstante et al 2009) Also slow winter precipitation rechargesater storage in deep soil horizons where it is protected from soil
vaporation The PWP increases the water stored in soil until a crit-cal level is reached that is required to break bud dormancy andence start tree growth This result is consistent with other results
rom nearby areas (Gonzaacutelez-Elizondo et al 2005)In regards to the negative contribution of CSP to growth at high
levations it could be explained by tree water stress In summerhese sites reach maximum soil water holding capacity affectingree growth due to the activity of soil microorganisms (Bartelt-yser et al 2005) Furthermore Li et al (2012) reported thatloudiness is an important factor that reduces growth through lightimitation
Tmax had a negative influence on TRI suggesting that high tem-eratures may lead to physiological impairment According toebourgeois et al (2012) the optimal photosynthesis rate corre-ponds to medium temperatures An acceleration of warming couldead to reduced tree vigor and increased risk of mortality or sus-eptibility to insect attack Tmax may indicate high vapor pressureeficit which cause the stomata to close thus reducing photosyn-hesis Allocation of carbohydrates is of major importance for leafnd root expansion which affects subsequent radial growth of treesMisson et al 2004)
These findings suggest that yearly growth in the high-elevationorest was limited more by maximum temperatures than meanemperatures This is consistent with findings from Adams and Kolb2005) whose research suggested that an increasing respirationate (ie carbon allocation to respiration) during periods of highemperatures could limit radial growth in temperate ecosystems
There was a strong positive correlation between Tmin and radialrowth There are a number of possible ecophysiological mecha-isms whereby a cold winter could be associated with tree growthinimal temperatures during winter will usually result in soil
reezing and a delay in growth until the ground has thawed in thepring (Chhin et al 2008) The positive response to Tmin is consis-ent with previous studies on other conifers in mountainous areasHuang et al 2010)
Along the whole elevational gradient correlation analysis sug-ested that warmer Tmin enhanced growth Tmin paired with PWPlays a major role as a predictor variable of tree growth par-icularly in higher-elevation stands where it explains a largeercentage of the variance in regression models These interac-ions contribute to prolonging the growing season of P cooperihe underlying mechanism of this effect could be interpreted toe that growth is enhanced by a low evapotranspiration rate inombination with water availability in the upper zones (Rossit al 2008) Together these environmental conditions promoten early budburst cambial reactivation and thus radial growthLebourgeois et al 2012) Lenoir et al (2008) has reported a similarrend
Tree growth at H LW L and HE zones was strongly associatedith climate whereas M zone had less-sensitive chronologies witho clear common response pattern Lower correlation between
rowth and climate variables particularly for lower and drierones has often been interpreted as a strategy to avoid ecophys-ological impairment during the driest period (Lebourgeois et al012)
KaTt
onologia 31 (2013) 198ndash 204 203
The elevational differences in the R2 and parameter regres-ion values did not reflect noteworthy elevational trends in radialrowth Indeed there were no discernible differences across eleva-ions in the radial growth pattern or climate growth relationshipsf P cooperi This is inconsistent with the concept of differing cli-atic signals at different elevations captured in tree ring data as
hown by Bickford et al (2011) Instead the differences we foundre probably due to specific site conditions The lack of system-tic trends across the elevational gradient is also due to the uniqueeography ecology and topography of the sites sampled (Tardift al 2003) in contrast with the ideal area for dendroclimato-ogical studies reported by Li et al (2012) Another explanationould be that trees responded to the climatic signals based onenetic adaptations throughout their range (Prior et al 2012) Itas been demonstrated that coniferous populations differentiateenetically in response to differential selection pressure occurringlong the elevational gradient The populations of lower altitudesave lower cold tolerance and higher growth potential (earlierhoot elongation later growth cessation and larger total shootlongation) than those of higher altitudes (Viveros-Viveros et al009) Dendrochronology approaches may have the potential toetect differences among populations to detect suitable genotypesor forest management strategies under climate change (Chen et al010) Stan and Daniels (2010) found that the complementarypplication of basal area improves the predictive capacity of treerowth rings in coastal British Columbia
onclusion
The results in this study did not indicate systematic trends alongn elevational gradient in tree growth response to climatic vari-tion in contrast with our hypothesis However some commonlimatic variables independent of local microclimate factors wererivers in regression models across the elevational zones Perhapsenetic adaptations have occurred as a response of populations toocal environmental variation
Radial tree growth was mainly correlated with previous climateonditions Tree growth was positively and significantly correlatedith previous winter conditions at most of the sites along the gra-ient Wet and cold winters were positively associated with radialrowth at most elevations whereas warm winters affected treerowth negatively The results showed that tree growth did notespond to precipitation or temperature as independent variables Pnd T were useful explanatory variables indicating the importancef future climate change which could significantly influence treerowth during the 21st century These projections have implica-ions for P cooperi radial growth For example a potential reductionn radial growth could be expected in response to less abundant
inter precipitation plus higher temperatures
cknowledgments
This work was conducted within the framework of Academicobility supported by the UJED amp CONACyT ldquoEstancias Posdoc-
orales y Sabaacuteticas al Extranjero para la Consolidacioacuten de Grupose InvestigacioacutenrdquoldquoPostdoctoral and Sabbatical Stays Abroad foronsolidation of Researchers Teamsrdquo We would like to thank theupport given by School of Forestry at Northern Arizona Univer-ity Also we recognize Larissa Yocom Amanda Stan Thomas E
olb and Jim Allen for their helpful assistance Omar Duraacuten Guerrand Joseacute Santiago Saacutenchez provided help with field data gatheringhe authors are grateful to Editors and anonymous reviewers forheir useful comments and suggestions
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)
rochr
T
mminuslaw
HSFntf
tt
wewmPaLvo
M Pompa-Garciacutea et al Dend
ree growthndashclimate relationships
As shown in Fig 3 TRI was significantly correlated with cli-ate over the study area with correlation coefficients ranging from039 to 046 The Pearsonrsquos coefficients (significance at the 005
evel) indicated that P and Tmin were positively correlated with TRIcross the elevational gradient Tmax showed a negative correlationith TRI for all elevations Tmean did not have a consistent trend
The relationship between TRI and P were similar in zones HE L and LW TRI was negatively influenced by previous Augusteptember and December temperature and current January andebruary Tmax temperatures for HE H and M but LW showed
egative correlations with March April Jun July and Septemberemperature In contrast an inconsistent pattern was observedor Tmean across elevational gradients Compared with other
f
t
Fig 3 Pearsonrsquos correlation coefficients for the TRI of Pinus cooperi against clim
onologia 31 (2013) 198ndash 204 201
emperature variables Tmin correlations were more consistent overhe entire gradient
Linear regression indicated that TRI was significantly associatedith climatic variables Table 3 shows variables specific to different
levations For TRI most of the variables selected for the modelsere those from the previous year plus CSP and CSTmin PWP wasainly associated with TRI (Pr F lt 00001) for all sites followed by
WTmax The stepwise regression did not find a consistent trendlong the gradient for dependent variable Models at the H HE LW and M sites explained 44 38 36 33 and 17 of the totalariance of TRI (respectively) The small mean square errors weref the H M and HE zones is consistent with small values computed
or the Mallowrsquos coefficient
The highest parameter estimates obtained for TRI with mul-ivariate models indicated that PWP had a greater association
ate variables from 1946 to 2010 (statistically significant values p lt 005)
202 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
Table 3Models for TRI selected by stepwise regression for different climate variables along elevational zones at the 010 significance level R2 Coefficient determination C(p) TheMallowrsquos coefficient
Altitude Variable Parameter estimate Standard error Mean square error R2 C(p) Pr gt F
Table 4Models for TRI selected by stepwise regression for climate variables along an elevational gradient at the 010 significance level R2 Coefficient determination
Altitude Variable Parameter estimate Standard error Mean square error R2 Pr gt F
ith growth in the L and HE zones Linear regression resultsemonstrated significant contributions by PWP and PWTmax tohe TRI models (Table 4) However neither showed a consistentrend across elevations These variables indicated that in this Pooperi population climate signals may explain temporal variationn TRI
iscussion
Our study provides the first accurately dated and statisticallyeliable records of P cooperi growth response to climatic vari-tion in Mexico Similar trends in growth were noted acrossve different elevations The statistics presented in Table 2 showhat TRI records contributes in understanding about relationshipsf regional weather on tree growth from mountain landscapeshat are characterized by gradual changes in precipitation andemperature (Adams and Kolb 2005) The values for mean inter-orrelation and mean sensitivity were similar to those reported forhronologies of conifers growing in northern Mexico (Gonzaacutelez-
lizondo et al 2005 Cleaveland et al 2003 Constante et al009) Most of the tree-ring chronologies had a moderate first orderutoregressive component indicating that previous year condi-ions had little effect on current year radial growth (Fritts 2001)
tmcd
lt0000106588
PS provided valuable information as an estimator of the qual-ty of the climatendashgrowth relationships assessment Meacuterian et al2013) reported that in naturally regenerated forests EPS could bexpected to decrease due to inter-tree differences in age andorize They also noted that coring two trees per plot instead of oneonsiderably reduced the noise of the regional chronology Evenhough some differences in TRI across the gradient were foundeneral trends in ring width variation were similar An increasederiod of growth preceded by a slow period of growth during the950s 1960s 1970s 1990s and 2000s was coincident with severeegional drought reported in nearby ecosystems (Diacuteaz et al 2002illanueva et al 2005 2009) These similarities suggest that Pooperi growth is influenced by regional climate signals Neverthe-ess there are some differences in climate responses between sitesue to specific site variation
Differences in sensitivity of tree radial growth were notedcross the elevational gradient from 2365 to 2905 masl Correlationnalysis for P cooperi showed that monthly P was most stronglyorrelated with radial growth across the whole gradient Precipi-
ation in previous winters was particularly important Tmin had a
ore significant contribution than Tmean whereas Tmax Pearsonrsquosoefficients were negatively associated with radial growth acrossifferent elevations
rochr
etswsrl(weihf
ettRcl
pLslcdta(
ft(rt
gnMfst(
gptptTbcea(t
wngzi2
sgtomsaagelcghgahse2df2ag
C
aacdgl
cwdggraogtiw
A
MtdCss
M Pompa-Garciacutea et al Dend
The radial growth of P cooperi at all elevations was mainly influ-nced by PWP Tables 3 and 4 suggest that water storage during theree dormancy period is useful as a reserve for the next growing sea-on (Litton et al 2007) This finding supports the hypothesis thatet winters contributed to subsequent warm-season photosynthe-
is and therefore to tree growth (Kerhoulas et al 2013) Winterains are usually of low intensity favoring high infiltration andow evapotranspiration which result in a positive water balanceConstante et al 2009) Also slow winter precipitation rechargesater storage in deep soil horizons where it is protected from soil
vaporation The PWP increases the water stored in soil until a crit-cal level is reached that is required to break bud dormancy andence start tree growth This result is consistent with other results
rom nearby areas (Gonzaacutelez-Elizondo et al 2005)In regards to the negative contribution of CSP to growth at high
levations it could be explained by tree water stress In summerhese sites reach maximum soil water holding capacity affectingree growth due to the activity of soil microorganisms (Bartelt-yser et al 2005) Furthermore Li et al (2012) reported thatloudiness is an important factor that reduces growth through lightimitation
Tmax had a negative influence on TRI suggesting that high tem-eratures may lead to physiological impairment According toebourgeois et al (2012) the optimal photosynthesis rate corre-ponds to medium temperatures An acceleration of warming couldead to reduced tree vigor and increased risk of mortality or sus-eptibility to insect attack Tmax may indicate high vapor pressureeficit which cause the stomata to close thus reducing photosyn-hesis Allocation of carbohydrates is of major importance for leafnd root expansion which affects subsequent radial growth of treesMisson et al 2004)
These findings suggest that yearly growth in the high-elevationorest was limited more by maximum temperatures than meanemperatures This is consistent with findings from Adams and Kolb2005) whose research suggested that an increasing respirationate (ie carbon allocation to respiration) during periods of highemperatures could limit radial growth in temperate ecosystems
There was a strong positive correlation between Tmin and radialrowth There are a number of possible ecophysiological mecha-isms whereby a cold winter could be associated with tree growthinimal temperatures during winter will usually result in soil
reezing and a delay in growth until the ground has thawed in thepring (Chhin et al 2008) The positive response to Tmin is consis-ent with previous studies on other conifers in mountainous areasHuang et al 2010)
Along the whole elevational gradient correlation analysis sug-ested that warmer Tmin enhanced growth Tmin paired with PWPlays a major role as a predictor variable of tree growth par-icularly in higher-elevation stands where it explains a largeercentage of the variance in regression models These interac-ions contribute to prolonging the growing season of P cooperihe underlying mechanism of this effect could be interpreted toe that growth is enhanced by a low evapotranspiration rate inombination with water availability in the upper zones (Rossit al 2008) Together these environmental conditions promoten early budburst cambial reactivation and thus radial growthLebourgeois et al 2012) Lenoir et al (2008) has reported a similarrend
Tree growth at H LW L and HE zones was strongly associatedith climate whereas M zone had less-sensitive chronologies witho clear common response pattern Lower correlation between
rowth and climate variables particularly for lower and drierones has often been interpreted as a strategy to avoid ecophys-ological impairment during the driest period (Lebourgeois et al012)
KaTt
onologia 31 (2013) 198ndash 204 203
The elevational differences in the R2 and parameter regres-ion values did not reflect noteworthy elevational trends in radialrowth Indeed there were no discernible differences across eleva-ions in the radial growth pattern or climate growth relationshipsf P cooperi This is inconsistent with the concept of differing cli-atic signals at different elevations captured in tree ring data as
hown by Bickford et al (2011) Instead the differences we foundre probably due to specific site conditions The lack of system-tic trends across the elevational gradient is also due to the uniqueeography ecology and topography of the sites sampled (Tardift al 2003) in contrast with the ideal area for dendroclimato-ogical studies reported by Li et al (2012) Another explanationould be that trees responded to the climatic signals based onenetic adaptations throughout their range (Prior et al 2012) Itas been demonstrated that coniferous populations differentiateenetically in response to differential selection pressure occurringlong the elevational gradient The populations of lower altitudesave lower cold tolerance and higher growth potential (earlierhoot elongation later growth cessation and larger total shootlongation) than those of higher altitudes (Viveros-Viveros et al009) Dendrochronology approaches may have the potential toetect differences among populations to detect suitable genotypesor forest management strategies under climate change (Chen et al010) Stan and Daniels (2010) found that the complementarypplication of basal area improves the predictive capacity of treerowth rings in coastal British Columbia
onclusion
The results in this study did not indicate systematic trends alongn elevational gradient in tree growth response to climatic vari-tion in contrast with our hypothesis However some commonlimatic variables independent of local microclimate factors wererivers in regression models across the elevational zones Perhapsenetic adaptations have occurred as a response of populations toocal environmental variation
Radial tree growth was mainly correlated with previous climateonditions Tree growth was positively and significantly correlatedith previous winter conditions at most of the sites along the gra-ient Wet and cold winters were positively associated with radialrowth at most elevations whereas warm winters affected treerowth negatively The results showed that tree growth did notespond to precipitation or temperature as independent variables Pnd T were useful explanatory variables indicating the importancef future climate change which could significantly influence treerowth during the 21st century These projections have implica-ions for P cooperi radial growth For example a potential reductionn radial growth could be expected in response to less abundant
inter precipitation plus higher temperatures
cknowledgments
This work was conducted within the framework of Academicobility supported by the UJED amp CONACyT ldquoEstancias Posdoc-
orales y Sabaacuteticas al Extranjero para la Consolidacioacuten de Grupose InvestigacioacutenrdquoldquoPostdoctoral and Sabbatical Stays Abroad foronsolidation of Researchers Teamsrdquo We would like to thank theupport given by School of Forestry at Northern Arizona Univer-ity Also we recognize Larissa Yocom Amanda Stan Thomas E
olb and Jim Allen for their helpful assistance Omar Duraacuten Guerrand Joseacute Santiago Saacutenchez provided help with field data gatheringhe authors are grateful to Editors and anonymous reviewers forheir useful comments and suggestions
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)
202 M Pompa-Garciacutea et al Dendrochronologia 31 (2013) 198ndash 204
Table 3Models for TRI selected by stepwise regression for different climate variables along elevational zones at the 010 significance level R2 Coefficient determination C(p) TheMallowrsquos coefficient
Altitude Variable Parameter estimate Standard error Mean square error R2 C(p) Pr gt F
Table 4Models for TRI selected by stepwise regression for climate variables along an elevational gradient at the 010 significance level R2 Coefficient determination
Altitude Variable Parameter estimate Standard error Mean square error R2 Pr gt F
ith growth in the L and HE zones Linear regression resultsemonstrated significant contributions by PWP and PWTmax tohe TRI models (Table 4) However neither showed a consistentrend across elevations These variables indicated that in this Pooperi population climate signals may explain temporal variationn TRI
iscussion
Our study provides the first accurately dated and statisticallyeliable records of P cooperi growth response to climatic vari-tion in Mexico Similar trends in growth were noted acrossve different elevations The statistics presented in Table 2 showhat TRI records contributes in understanding about relationshipsf regional weather on tree growth from mountain landscapeshat are characterized by gradual changes in precipitation andemperature (Adams and Kolb 2005) The values for mean inter-orrelation and mean sensitivity were similar to those reported forhronologies of conifers growing in northern Mexico (Gonzaacutelez-
lizondo et al 2005 Cleaveland et al 2003 Constante et al009) Most of the tree-ring chronologies had a moderate first orderutoregressive component indicating that previous year condi-ions had little effect on current year radial growth (Fritts 2001)
tmcd
lt0000106588
PS provided valuable information as an estimator of the qual-ty of the climatendashgrowth relationships assessment Meacuterian et al2013) reported that in naturally regenerated forests EPS could bexpected to decrease due to inter-tree differences in age andorize They also noted that coring two trees per plot instead of oneonsiderably reduced the noise of the regional chronology Evenhough some differences in TRI across the gradient were foundeneral trends in ring width variation were similar An increasederiod of growth preceded by a slow period of growth during the950s 1960s 1970s 1990s and 2000s was coincident with severeegional drought reported in nearby ecosystems (Diacuteaz et al 2002illanueva et al 2005 2009) These similarities suggest that Pooperi growth is influenced by regional climate signals Neverthe-ess there are some differences in climate responses between sitesue to specific site variation
Differences in sensitivity of tree radial growth were notedcross the elevational gradient from 2365 to 2905 masl Correlationnalysis for P cooperi showed that monthly P was most stronglyorrelated with radial growth across the whole gradient Precipi-
ation in previous winters was particularly important Tmin had a
ore significant contribution than Tmean whereas Tmax Pearsonrsquosoefficients were negatively associated with radial growth acrossifferent elevations
rochr
etswsrl(weihf
ettRcl
pLslcdta(
ft(rt
gnMfst(
gptptTbcea(t
wngzi2
sgtomsaagelcghgahse2df2ag
C
aacdgl
cwdggraogtiw
A
MtdCss
M Pompa-Garciacutea et al Dend
The radial growth of P cooperi at all elevations was mainly influ-nced by PWP Tables 3 and 4 suggest that water storage during theree dormancy period is useful as a reserve for the next growing sea-on (Litton et al 2007) This finding supports the hypothesis thatet winters contributed to subsequent warm-season photosynthe-
is and therefore to tree growth (Kerhoulas et al 2013) Winterains are usually of low intensity favoring high infiltration andow evapotranspiration which result in a positive water balanceConstante et al 2009) Also slow winter precipitation rechargesater storage in deep soil horizons where it is protected from soil
vaporation The PWP increases the water stored in soil until a crit-cal level is reached that is required to break bud dormancy andence start tree growth This result is consistent with other results
rom nearby areas (Gonzaacutelez-Elizondo et al 2005)In regards to the negative contribution of CSP to growth at high
levations it could be explained by tree water stress In summerhese sites reach maximum soil water holding capacity affectingree growth due to the activity of soil microorganisms (Bartelt-yser et al 2005) Furthermore Li et al (2012) reported thatloudiness is an important factor that reduces growth through lightimitation
Tmax had a negative influence on TRI suggesting that high tem-eratures may lead to physiological impairment According toebourgeois et al (2012) the optimal photosynthesis rate corre-ponds to medium temperatures An acceleration of warming couldead to reduced tree vigor and increased risk of mortality or sus-eptibility to insect attack Tmax may indicate high vapor pressureeficit which cause the stomata to close thus reducing photosyn-hesis Allocation of carbohydrates is of major importance for leafnd root expansion which affects subsequent radial growth of treesMisson et al 2004)
These findings suggest that yearly growth in the high-elevationorest was limited more by maximum temperatures than meanemperatures This is consistent with findings from Adams and Kolb2005) whose research suggested that an increasing respirationate (ie carbon allocation to respiration) during periods of highemperatures could limit radial growth in temperate ecosystems
There was a strong positive correlation between Tmin and radialrowth There are a number of possible ecophysiological mecha-isms whereby a cold winter could be associated with tree growthinimal temperatures during winter will usually result in soil
reezing and a delay in growth until the ground has thawed in thepring (Chhin et al 2008) The positive response to Tmin is consis-ent with previous studies on other conifers in mountainous areasHuang et al 2010)
Along the whole elevational gradient correlation analysis sug-ested that warmer Tmin enhanced growth Tmin paired with PWPlays a major role as a predictor variable of tree growth par-icularly in higher-elevation stands where it explains a largeercentage of the variance in regression models These interac-ions contribute to prolonging the growing season of P cooperihe underlying mechanism of this effect could be interpreted toe that growth is enhanced by a low evapotranspiration rate inombination with water availability in the upper zones (Rossit al 2008) Together these environmental conditions promoten early budburst cambial reactivation and thus radial growthLebourgeois et al 2012) Lenoir et al (2008) has reported a similarrend
Tree growth at H LW L and HE zones was strongly associatedith climate whereas M zone had less-sensitive chronologies witho clear common response pattern Lower correlation between
rowth and climate variables particularly for lower and drierones has often been interpreted as a strategy to avoid ecophys-ological impairment during the driest period (Lebourgeois et al012)
KaTt
onologia 31 (2013) 198ndash 204 203
The elevational differences in the R2 and parameter regres-ion values did not reflect noteworthy elevational trends in radialrowth Indeed there were no discernible differences across eleva-ions in the radial growth pattern or climate growth relationshipsf P cooperi This is inconsistent with the concept of differing cli-atic signals at different elevations captured in tree ring data as
hown by Bickford et al (2011) Instead the differences we foundre probably due to specific site conditions The lack of system-tic trends across the elevational gradient is also due to the uniqueeography ecology and topography of the sites sampled (Tardift al 2003) in contrast with the ideal area for dendroclimato-ogical studies reported by Li et al (2012) Another explanationould be that trees responded to the climatic signals based onenetic adaptations throughout their range (Prior et al 2012) Itas been demonstrated that coniferous populations differentiateenetically in response to differential selection pressure occurringlong the elevational gradient The populations of lower altitudesave lower cold tolerance and higher growth potential (earlierhoot elongation later growth cessation and larger total shootlongation) than those of higher altitudes (Viveros-Viveros et al009) Dendrochronology approaches may have the potential toetect differences among populations to detect suitable genotypesor forest management strategies under climate change (Chen et al010) Stan and Daniels (2010) found that the complementarypplication of basal area improves the predictive capacity of treerowth rings in coastal British Columbia
onclusion
The results in this study did not indicate systematic trends alongn elevational gradient in tree growth response to climatic vari-tion in contrast with our hypothesis However some commonlimatic variables independent of local microclimate factors wererivers in regression models across the elevational zones Perhapsenetic adaptations have occurred as a response of populations toocal environmental variation
Radial tree growth was mainly correlated with previous climateonditions Tree growth was positively and significantly correlatedith previous winter conditions at most of the sites along the gra-ient Wet and cold winters were positively associated with radialrowth at most elevations whereas warm winters affected treerowth negatively The results showed that tree growth did notespond to precipitation or temperature as independent variables Pnd T were useful explanatory variables indicating the importancef future climate change which could significantly influence treerowth during the 21st century These projections have implica-ions for P cooperi radial growth For example a potential reductionn radial growth could be expected in response to less abundant
inter precipitation plus higher temperatures
cknowledgments
This work was conducted within the framework of Academicobility supported by the UJED amp CONACyT ldquoEstancias Posdoc-
orales y Sabaacuteticas al Extranjero para la Consolidacioacuten de Grupose InvestigacioacutenrdquoldquoPostdoctoral and Sabbatical Stays Abroad foronsolidation of Researchers Teamsrdquo We would like to thank theupport given by School of Forestry at Northern Arizona Univer-ity Also we recognize Larissa Yocom Amanda Stan Thomas E
olb and Jim Allen for their helpful assistance Omar Duraacuten Guerrand Joseacute Santiago Saacutenchez provided help with field data gatheringhe authors are grateful to Editors and anonymous reviewers forheir useful comments and suggestions
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)
rochr
etswsrl(weihf
ettRcl
pLslcdta(
ft(rt
gnMfst(
gptptTbcea(t
wngzi2
sgtomsaagelcghgahse2df2ag
C
aacdgl
cwdggraogtiw
A
MtdCss
M Pompa-Garciacutea et al Dend
The radial growth of P cooperi at all elevations was mainly influ-nced by PWP Tables 3 and 4 suggest that water storage during theree dormancy period is useful as a reserve for the next growing sea-on (Litton et al 2007) This finding supports the hypothesis thatet winters contributed to subsequent warm-season photosynthe-
is and therefore to tree growth (Kerhoulas et al 2013) Winterains are usually of low intensity favoring high infiltration andow evapotranspiration which result in a positive water balanceConstante et al 2009) Also slow winter precipitation rechargesater storage in deep soil horizons where it is protected from soil
vaporation The PWP increases the water stored in soil until a crit-cal level is reached that is required to break bud dormancy andence start tree growth This result is consistent with other results
rom nearby areas (Gonzaacutelez-Elizondo et al 2005)In regards to the negative contribution of CSP to growth at high
levations it could be explained by tree water stress In summerhese sites reach maximum soil water holding capacity affectingree growth due to the activity of soil microorganisms (Bartelt-yser et al 2005) Furthermore Li et al (2012) reported thatloudiness is an important factor that reduces growth through lightimitation
Tmax had a negative influence on TRI suggesting that high tem-eratures may lead to physiological impairment According toebourgeois et al (2012) the optimal photosynthesis rate corre-ponds to medium temperatures An acceleration of warming couldead to reduced tree vigor and increased risk of mortality or sus-eptibility to insect attack Tmax may indicate high vapor pressureeficit which cause the stomata to close thus reducing photosyn-hesis Allocation of carbohydrates is of major importance for leafnd root expansion which affects subsequent radial growth of treesMisson et al 2004)
These findings suggest that yearly growth in the high-elevationorest was limited more by maximum temperatures than meanemperatures This is consistent with findings from Adams and Kolb2005) whose research suggested that an increasing respirationate (ie carbon allocation to respiration) during periods of highemperatures could limit radial growth in temperate ecosystems
There was a strong positive correlation between Tmin and radialrowth There are a number of possible ecophysiological mecha-isms whereby a cold winter could be associated with tree growthinimal temperatures during winter will usually result in soil
reezing and a delay in growth until the ground has thawed in thepring (Chhin et al 2008) The positive response to Tmin is consis-ent with previous studies on other conifers in mountainous areasHuang et al 2010)
Along the whole elevational gradient correlation analysis sug-ested that warmer Tmin enhanced growth Tmin paired with PWPlays a major role as a predictor variable of tree growth par-icularly in higher-elevation stands where it explains a largeercentage of the variance in regression models These interac-ions contribute to prolonging the growing season of P cooperihe underlying mechanism of this effect could be interpreted toe that growth is enhanced by a low evapotranspiration rate inombination with water availability in the upper zones (Rossit al 2008) Together these environmental conditions promoten early budburst cambial reactivation and thus radial growthLebourgeois et al 2012) Lenoir et al (2008) has reported a similarrend
Tree growth at H LW L and HE zones was strongly associatedith climate whereas M zone had less-sensitive chronologies witho clear common response pattern Lower correlation between
rowth and climate variables particularly for lower and drierones has often been interpreted as a strategy to avoid ecophys-ological impairment during the driest period (Lebourgeois et al012)
KaTt
onologia 31 (2013) 198ndash 204 203
The elevational differences in the R2 and parameter regres-ion values did not reflect noteworthy elevational trends in radialrowth Indeed there were no discernible differences across eleva-ions in the radial growth pattern or climate growth relationshipsf P cooperi This is inconsistent with the concept of differing cli-atic signals at different elevations captured in tree ring data as
hown by Bickford et al (2011) Instead the differences we foundre probably due to specific site conditions The lack of system-tic trends across the elevational gradient is also due to the uniqueeography ecology and topography of the sites sampled (Tardift al 2003) in contrast with the ideal area for dendroclimato-ogical studies reported by Li et al (2012) Another explanationould be that trees responded to the climatic signals based onenetic adaptations throughout their range (Prior et al 2012) Itas been demonstrated that coniferous populations differentiateenetically in response to differential selection pressure occurringlong the elevational gradient The populations of lower altitudesave lower cold tolerance and higher growth potential (earlierhoot elongation later growth cessation and larger total shootlongation) than those of higher altitudes (Viveros-Viveros et al009) Dendrochronology approaches may have the potential toetect differences among populations to detect suitable genotypesor forest management strategies under climate change (Chen et al010) Stan and Daniels (2010) found that the complementarypplication of basal area improves the predictive capacity of treerowth rings in coastal British Columbia
onclusion
The results in this study did not indicate systematic trends alongn elevational gradient in tree growth response to climatic vari-tion in contrast with our hypothesis However some commonlimatic variables independent of local microclimate factors wererivers in regression models across the elevational zones Perhapsenetic adaptations have occurred as a response of populations toocal environmental variation
Radial tree growth was mainly correlated with previous climateonditions Tree growth was positively and significantly correlatedith previous winter conditions at most of the sites along the gra-ient Wet and cold winters were positively associated with radialrowth at most elevations whereas warm winters affected treerowth negatively The results showed that tree growth did notespond to precipitation or temperature as independent variables Pnd T were useful explanatory variables indicating the importancef future climate change which could significantly influence treerowth during the 21st century These projections have implica-ions for P cooperi radial growth For example a potential reductionn radial growth could be expected in response to less abundant
inter precipitation plus higher temperatures
cknowledgments
This work was conducted within the framework of Academicobility supported by the UJED amp CONACyT ldquoEstancias Posdoc-
orales y Sabaacuteticas al Extranjero para la Consolidacioacuten de Grupose InvestigacioacutenrdquoldquoPostdoctoral and Sabbatical Stays Abroad foronsolidation of Researchers Teamsrdquo We would like to thank theupport given by School of Forestry at Northern Arizona Univer-ity Also we recognize Larissa Yocom Amanda Stan Thomas E
olb and Jim Allen for their helpful assistance Omar Duraacuten Guerrand Joseacute Santiago Saacutenchez provided help with field data gatheringhe authors are grateful to Editors and anonymous reviewers forheir useful comments and suggestions
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)
2 rochr
R
A
B
B
B
B
C
C
C
C
C
C
C
C
C
D
FG
G
H
H
K
L
L
L
L
L
MM
M
P
R
S
S
S
S
T
V
V
V
W
W
Z
Web references
04 M Pompa-Garciacutea et al Dend
eferences
dams HD Kolb TE 2005 Tree growth response to drought and temperature ina mountain landscape in northern Arizona USA Journal of Biogeography 321629ndash1640 httpdxdoiorg101111j1365-2699200501292x
artelt-Ryser J Joshi J Schmid B Brandl H Balser T 2005 Soil feed-backs of plant diversity on soil microbial communities and subsequent plantgrowth Perspectives in Plant Ecology Evolution and Systematics 7 27ndash49httpdxdoiorg101016jppees200411002
eniston M 2002 Climate modeling at various spatial and temporal scales wherecan dendrochronology help Dendrochronologia 20 117ndash131
ickford IN Fuleacute ZP Kolb TE 2011 Growth sensitivity to drought of co-occurring Pinus spp along an elevation gradient in northern Mexico WesternNorth American Naturalist 71 (3) 338ndash348
iondi F Waikul J 2004 DENDROCLIM2002 a C++ program for statistical calibra-tion of climate signals in tree-ring chronologies Computers amp Geosciences 30303ndash311 httpdxdoiorg101016jcageo200311004
andel-Peacuterez D Linares JC Vinegla B Lucas-Borja ME 2012 Assessingclimatendashgrowth relationships under contrasting stands of co-occurring Iberianpines along an altitudinal gradient Forest Ecology and Management 274 48ndash57httpdxdoiorg101016jforeco201202010
hen P Welsh C Hamann A 2010 Geographic variation in growthresponse of Douglas-fir to inter-annual climate variability andprojected climate change Global Change Biology 16 3374ndash3385httpdxdoiorg101111j1365-2486201002166x
hhin S Hogg EH Lieffers VJ Huang S 2008 Influences of climateon the radial growth of lodgepole pine in Alberta Botany 86 167ndash178httpdxdoiorg101139B07-120
ook ER Holmes RH 1984 Program ARSTAN and Users Manual Laboratory ofTree-Ring Research University of Arizona Tucson AZ USA 15 p
ook ER 1987 The decomposition of tree-ring series for environmental studiesTree-Ring Bulletin 47 37ndash59
onstante GV Villanueva DJ Cerano PJ Cornejo OEH Valencia MS 2009Dendrocronologiacutea de Pinus cembroides Zucc y reconstruccioacuten de precipitacioacutenestacional para el sureste de Coahuila Revista Ciencia Forestal en Meacutexico 3417ndash38
NA (Comisioacuten Nacional del Agua) 2012 Datos climaacuteticos de estaciones meteo-roloacutegicas de Durango El Salto
ruz CF de los Santos PHM Valdez LJR 2008 Sistema compatible deahusamiento-volumen para Pinus cooperi Blanco en Durango Meacutexico Agrocien-cia 42 (4) 473ndash485
iacuteaz SC Therrell M Stahle DW Cleaveland MK 2002 Chihuahuawinterndashspring precipitation reconstructed from tree-rings 1647ndash1992 ClimateResearch 22 237ndash244
ritts HC 2001 Tree Rings and Climate Blackburn Press Caldwell NJ 567 ponzaacutelez-Elizondo M Jurado E Naacutevar J Gonzaacutelez-Elizondo MS Villanueva
J Aguirre O Jimeacutenez J 2005 Tree-rings and climate relationships forDouglas-fir chronologies from the Sierra Madre Occidental Mexico a1681ndash2001 rain reconstruction Forest Ecology and Management 213 39ndash53httpdxdoiorg101016jforeco200503012
onzaacutelez-Elizondo MS Gonzaacutelez-Elizondo M Maacuterquez LMA 2007 Vegetacioacuteny ecorregiones de Durango Plaza y Valdeacutes SA de CV Meacutexico DF 219 p
olmes RL 1983 Computer-assisted quality control in tree-ring dating and mea-surement Tree-Ring Bulletin 43 69ndash78
uang J Tardif JC Bergeron Y Denneler B Berninger F Girardin MP 2010Radial growth response of four dominant boreal tree species to climate along alatitudinal gradient in the eastern Canadian boreal forest Global Change Biology16 711ndash731 httpdxdoiorg101111j1365-2486200901990x
erhoulas LP Kolb TE Koch GW 2013 Tree size stand densityand the source of water used across seasons by ponderosa pine innorthern Arizona Forest Ecology and Management 289 425ndash433httpdxdoiorg101016jforeco201210036
ebourgeois F Meacuterian P Courdier F Ladier J Dreyfus P 2012 Insta-bility of climate signal in tree-ring width in Mediterranean mountainsa multi-species analysis Trees Structure and Function 26 715ndash729httpdxdoiorg101007s00468-011-0638-7
enoir J Geacutegout JC Marquet PA de Ruffray P Brisse H 2008 A significantupward shift in plant species optimum elevation during the 20th century Sci-ence 320 1768ndash1771 httpdxdoiorg101126science1156831
i ZS Liu GH Fu BJ Hu CJ Luo SZ Liu XL He F 2012 Anomaloustemperaturendashgrowth response of Abies faxoniana to sustained freezing stress
P
onologia 31 (2013) 198ndash 204
along elevational gradients in Chinarsquos Western Sichuan Province Trees 261373ndash1388 httpdxdoiorg101007s00468-012-0712-9
itton CM Raich JW Ryan MG 2007 Carbon allocation inforest ecosystems Global Change Biology 13 2089ndash2109httpdxdoiorg101111j1365-2486200701420x
o YH Blanco JA Seely B Welham C Kimmins JP 2010 Rela-tionships between climate and tree radial growth in interior BritishColumbia Canada Forest Ecology and Management 259 932ndash942httpdxdoiorg101016jforeco200911033
allows CL 1973 Some comments on Cp Technometrics 15 661ndash675eacuterian P Pierrat JC Lebourgeois F 2013 Effect of sampling effort on the
regional chronology statistics and climatendashgrowth relationships estimationDendrochronologia 31 58ndash67 httpdxdoiorg101016jdendro201207001
isson L Rathgeber C Guiot J 2004 Dendroecological analysis of climaticeffects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model Canadian Journal of Forest Research 34 888ndash898httpdxdoiorg101139X03-253
rior LD Grierson PF McCaw WL Tng DYP Nichols SC Bowman DJMS2012 Variation in stem radial growth of the Australian conifer Callitris columel-laris across the worldrsquos driest and least fertile vegetated continent Trees 261169ndash1179 httpdxdoiorg101007s00468-012-0693-8
ossi S Deslauriers A Gric ar J Seo J-W Rathgeber CB Anfodillo T MorinH Levanic T Oven P Jalkanen R 2008 Critical temperatures for xylogene-sis in conifers of cold climates Global Ecology and Biogeography 17 696ndash707httpdxdoiorg101111j1466-8238200800417x
AS Institute 2004 SASSTAT Userrsquos Guide Release 91 SAS Institute Inc Cary NCUSA 2170 p
eager R Ting M Held I Kushnir Y Lu J Vecchi G Huang H-P Harnik NLeetmaa A Lau N-C Li C Velez J Naik N 2007 Model projections of animminent transition to a more arid climate in southwestern North AmericaScience 316 1181ndash1184 httpdxdoiorg101126science1139601
tan AB Daniels LD 2010 Calibrating the radial-growth aver-aging method for detecting releases in old-growth forests ofcoastal British Columbia Canada Dendrochronologia 28 135ndash147httpdxdoiorg101016jdendro200910003
tokes MA Smiley TL 1968 An Introduction to Tree-ring Dating University ofChicago Press Chicago USA 73 p
ardif J Camarero JJ Ribas M Gutieacuterrez E 2003 Spatiotem-poral variability in tree growth in the Central Pyreneesclimatic and site influences Ecological Monographs 73 241ndash257httpdxdoiorg1018900012-9615(2003)073[0241SVITGI]20CO2
illanueva DJ Luckman BH Stahle DW Therrell MD Cleaveland MKCerano PJ Gutieacuterrez GG Estrada AJ Jasso IR 2005 Hydroclimaticvariability of the upper Nazas basin water management implications forthe irrigated area of the Comarca Lagunera Dendrocronologia 22 215ndash223httpdxdoiorg101016jdendro200504005
illanueva DJ Fuleacute PZ Cerano PJ Estrada AJ Saacutenchez CI 2009 Recon-struccioacuten de la precipitacioacuten estacional para el barlovento de la Sierra MadreOccidental Revista Ciencia Forestal en Meacutexico 34 (105) 37ndash69
iveros-Viveros H Saenz-Romero C Vargas-Hernandez J Lopez-UptonJ Ramirez-Valverde G Santacruz-Varela A 2009 Altitudinal variationin Pinus hartwegii Lindl I height growth shoot phenology and frostdamage in seedlings Forest Ecology and Management 257 836ndash842httpdxdoiorg101016jforeco200810021
illiams AP Allen CD Macalady AK Griffin D Woodhouse CA Meko DMSwetnam TW Rauscher SA Seager R Grissino-Mayer HD Dean JS CookER Gangodagamage C Cai M McDowell NG 2012 Temperature as a potentdriver of regional forest drought stress and tree mortality Nature ClimateChange httpdxdoiorg101038NCLIMATE1693 (in press)
ilmking M Juday G Barber V Zald H 2004 Recent climate warmingforces contrasting growth responses of white spruce at treeline in Alaskathrough temperature thresholds Global Change Biology 10 1724ndash1736httpdxdoiorg101111j1365-2486200400826
hang QB Hebda RJ 2004 Variation in radial growth patterns of Pseudotsugamenziesii on the central coast of British Columbia Canada Canadian Journal ofForest Research 34 1946ndash1954 httpdxdoiorg101139X04-078
ompa GM 2012 Geographic Information Systems for Durangorsquos Ecosys-tems Available online httpwwwujedmxsigeedInicioaspx (retrieved on300912)