Latitudinal and Elevational Range Shifts under Contemporary Climate Change Jonathan Lenoir and Jens-Christian Svenning, Aarhus University, Aarhus, Denmark r 2013 Elsevier Inc. All rights reserved. Glossary Front edge Stable margin of a species’ range where population size is growing. Leading edge Dynamic margin of a species’ range where it is expanding into neighboring unoccupied areas. Range core Central part of the distribution of a species. Range periphery Marginal parts of the distribution of a species. Rear edge Stable margin of a species’ range where population size is declining. Species’ niche The scope of environmental conditions under which a species is able to maintain stable or increasing populations. Species’ optimum The value of a given environmental variable at which a species reaches its maximum probability of occurrence or abundance. Species’ range The spatial distribution of a species, that is, the sum of all geographic locations of its individuals over a given period in time. Species’ response curve The relationship between a species’ probability of occurrence or its density (number of individuals per unit area) and the values of a given environmental variable. Trailing edge Dynamic margin of a species’ range where it is becoming locally extinct, causing the range to retract. The concept of range shifts refers to the dynamic process whereby species shift their distribution over time, tracking geographic shifts in the climate and habitat conditions that they require. Range shifts are also commonly defined as migration events that occur on decadal to millennial or longer time scales, usually beyond the life span of individuals. In contrast, the concept does not include migration events occurring over the life span of individuals, that is, on a seasonal to annual or shorter time scales, as a result of recurring movements of individuals between spatially separate habitats in order to avoid temporarily unfavorable conditions. It also does not include essentially stochastic long-distance dispersal events of a single or few individuals that fail to establish new popu- lations. Recently, species range shifts have attracted a great deal of interest from the scientific community as a direct con- sequence of the increasing focus on current global environ- mental changes, notably the changing climate (IPCC, 2007a). There is now a growing body of studies reporting and fore- casting species range shifts across the main geographic dimen- sions (latitude, longitude, and elevation) as a response to both contemporary and future changes in climatic conditions. Here, the focus is on realized range shifts in response to contemporary climate change, a phenomenon that is of key importance for developing well-founded predictive models of range shifts under expected substantial future climate change (IPCC, 2007b). Because dramatic range shifts have also occurred in the geological past, species range shifts, until recently, have been described in textbooks through paleontological examples, but contemporary range shifts are now becoming even more evident. Species’ Range: Geographic Dimensions and Ecological Drivers As a species’ range can be defined by the geographic locations of all its individuals over a given period of time (Brown and Lomolino, 1998), it is often difficult and challenging to cap- ture and represent it. Several abstractions of the real distri- bution of a single species have been used, with the most frequent approaches being to represent a species’ range as outline, dot, or contour maps in a two-dimensional geo- graphic space, most often across the latitudinal and longi- tudinal dimensions. One could also represent a species’ range in a three-dimensional geographic space by adding occurrence along elevational gradients to the representation (Figure 1). Another more simplistic abstraction is to project the known or estimated occurrences of the species against one of these three dimensions. Hence, latitude, longitude, and elevation consti- tute the geographic dimensions over which a species’ range is estimated, and a species’ range shift is defined as changes in these geographic dimensions over the fourth dimension: time (Figure 1). A species’ range is generally understood to be at least partly determined by the species’ environmental requirements and tolerances, that is, its ecological niche (Hutchinson, 1957; Soberon and Nakamura, 2009). The ecological niche is typi- cally considered a multidimensional space (or ‘‘hypervo- lume’’), in which the different dimensions represent different environmental conditions (Hutchinson, 1957). Especially at large scales, climate is often thought to constitute the niche axes that are most relevant for species ranges (Pearson and Dawson, 2003). Therefore, one should be able to deduce a species’ climatic niche from its range. Unfortunately, the geographic distribution of a species is unlikely to fully repre- sent its ecological niche for at least three reasons (Colwell and Rangel, 2009): (1) geographic unavailability of some portions of its ecological niche; (2) geographic inaccessibility due to its limited dispersal abilities (Svenning and Skov, 2004; Blach- Overgaard et al., 2010); and (3) geographic control by con- straining species interactions (Case et al., 2005). Throughout this article, therefore, the species’ range is solely being con- sidered as an incomplete geographic reflection of the species’ niche. Still, it is useful to relate ranges to the niche concept in Encyclopedia of Biodiversity, Volume 4 http://dx.doi.org/10.1016/B978-0-12-384719-5.00375-0 599 Author's personal copy
Transcript
Latitudinal and Elevational Range Shifts under Contemporary Climate Change
Jonathan Lenoir and Jens-Christian Svenning, Aarhus University, Aarhus, Denmark
r 2013 Elsevier Inc. All rights reserved.
GlossaryFront edge Stable margin of a species’ range where
population size is growing.
Leading edge Dynamic margin of a species’ range where it
is expanding into neighboring unoccupied areas.
Range core Central part of the distribution of a species.
Range periphery Marginal parts of the distribution of a
species.
Rear edge Stable margin of a species’ range where
population size is declining.
Species’ niche The scope of environmental conditions
under which a species is able to maintain stable or
increasing populations.
Species’ optimum The value of a given environmental
variable at which a species reaches its maximum probability
of occurrence or abundance.
Species’ range The spatial distribution of a species, that is,
the sum of all geographic locations of its individuals over a
given period in time.
Species’ response curve The relationship between a
species’ probability of occurrence or its density (number of
individuals per unit area) and the values of a given
environmental variable.
Trailing edge Dynamic margin of a species’ range where
it is becoming locally extinct, causing the range to
retract.
The concept of range shifts refers to the dynamic process
whereby species shift their distribution over time, tracking
geographic shifts in the climate and habitat conditions that they
require. Range shifts are also commonly defined as migration
events that occur on decadal to millennial or longer time
scales, usually beyond the life span of individuals. In contrast,
the concept does not include migration events occurring
over the life span of individuals, that is, on a seasonal to annual
or shorter time scales, as a result of recurring movements
of individuals between spatially separate habitats in order
to avoid temporarily unfavorable conditions. It also does
not include essentially stochastic long-distance dispersal events
of a single or few individuals that fail to establish new popu-
lations. Recently, species range shifts have attracted a great deal
of interest from the scientific community as a direct con-
sequence of the increasing focus on current global environ-
mental changes, notably the changing climate (IPCC, 2007a).
There is now a growing body of studies reporting and fore-
casting species range shifts across the main geographic dimen-
sions (latitude, longitude, and elevation) as a response to both
contemporary and future changes in climatic conditions. Here,
the focus is on realized range shifts in response to contemporary
climate change, a phenomenon that is of key importance for
developing well-founded predictive models of range shifts
under expected substantial future climate change (IPCC,
2007b). Because dramatic range shifts have also occurred in the
geological past, species range shifts, until recently, have been
described in textbooks through paleontological examples, but
contemporary range shifts are now becoming even more
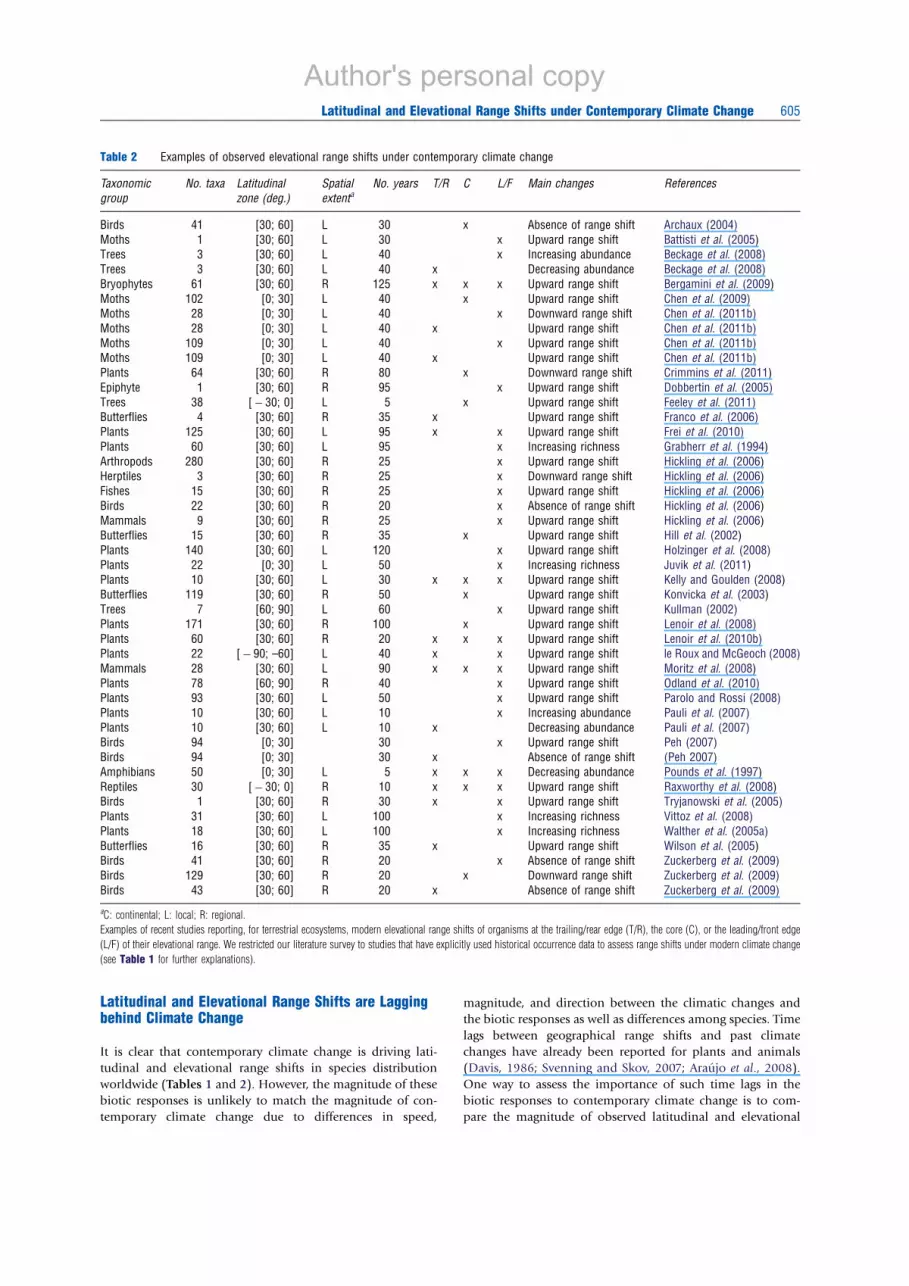
Arctic, and alpine) with much less research on elevational
range shifts in hotter areas such as Mediterranean and tropical
biomes (Table 2). However, on the contrary to the latitudinal
range shifts, there is relatively many studies documenting
elevational range shifts in the Tropics, for example, for plants,
insects, amphibians, and reptiles (Pounds et al., 1997; Rax-
worthy et al., 2008; Chen et al., 2009; Feeley et al., 2011; Juvik
et al., 2011). Continental-scale studies of elevational range
shifts under contemporary climate change are still lacking
(Table 2).
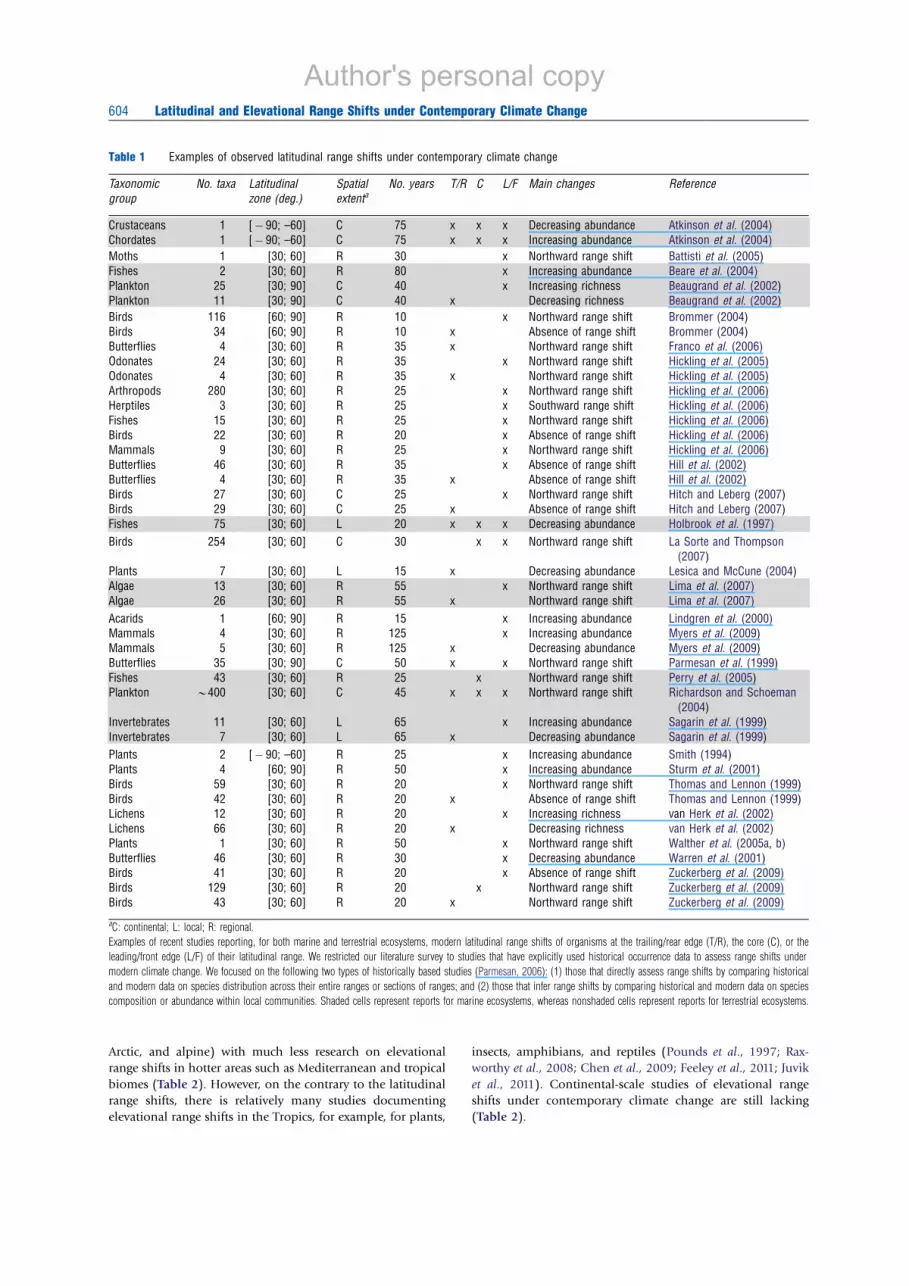
Table 1 Examples of observed latitudinal range shifts under contemporary climate change
Taxonomic
group
No. taxa Latitudinal
zone (deg.)
Spatial
extentaNo. years T/R C L/F Main changes Reference
Crustaceans 1 [ÿ 90; –60] C 75 x x x Decreasing abundance Atkinson et al. (2004)
Chordates 1 [ÿ 90; –60] C 75 x x x Increasing abundance Atkinson et al. (2004)
Moths 1 [30; 60] R 30 x Northward range shift Battisti et al. (2005)
Fishes 2 [30; 60] R 80 x Increasing abundance Beare et al. (2004)
Plankton 25 [30; 90] C 40 x Increasing richness Beaugrand et al. (2002)
Plankton 11 [30; 90] C 40 x Decreasing richness Beaugrand et al. (2002)
Birds 116 [60; 90] R 10 x Northward range shift Brommer (2004)
Birds 34 [60; 90] R 10 x Absence of range shift Brommer (2004)
Butterflies 4 [30; 60] R 35 x Northward range shift Franco et al. (2006)
Odonates 24 [30; 60] R 35 x Northward range shift Hickling et al. (2005)
Odonates 4 [30; 60] R 35 x Northward range shift Hickling et al. (2005)
Arthropods 280 [30; 60] R 25 x Northward range shift Hickling et al. (2006)
Herptiles 3 [30; 60] R 25 x Southward range shift Hickling et al. (2006)
Fishes 15 [30; 60] R 25 x Northward range shift Hickling et al. (2006)
Birds 22 [30; 60] R 20 x Absence of range shift Hickling et al. (2006)
Mammals 9 [30; 60] R 25 x Northward range shift Hickling et al. (2006)
Butterflies 46 [30; 60] R 35 x Absence of range shift Hill et al. (2002)
Butterflies 4 [30; 60] R 35 x Absence of range shift Hill et al. (2002)
Birds 27 [30; 60] C 25 x Northward range shift Hitch and Leberg (2007)
Birds 29 [30; 60] C 25 x Absence of range shift Hitch and Leberg (2007)
Fishes 75 [30; 60] L 20 x x x Decreasing abundance Holbrook et al. (1997)
Birds 254 [30; 60] C 30 x x Northward range shift La Sorte and Thompson
(2007)
Plants 7 [30; 60] L 15 x Decreasing abundance Lesica and McCune (2004)
Algae 13 [30; 60] R 55 x Northward range shift Lima et al. (2007)
Algae 26 [30; 60] R 55 x Northward range shift Lima et al. (2007)
Acarids 1 [60; 90] R 15 x Increasing abundance Lindgren et al. (2000)
Mammals 4 [30; 60] R 125 x Increasing abundance Myers et al. (2009)
Mammals 5 [30; 60] R 125 x Decreasing abundance Myers et al. (2009)
Butterflies 35 [30; 90] C 50 x x Northward range shift Parmesan et al. (1999)
Fishes 43 [30; 60] R 25 x Northward range shift Perry et al. (2005)
Plankton B400 [30; 60] C 45 x x x Northward range shift Richardson and Schoeman
(2004)
Invertebrates 11 [30; 60] L 65 x Increasing abundance Sagarin et al. (1999)
Invertebrates 7 [30; 60] L 65 x Decreasing abundance Sagarin et al. (1999)
Plants 2 [ÿ 90; –60] R 25 x Increasing abundance Smith (1994)
Plants 4 [60; 90] R 50 x Increasing abundance Sturm et al. (2001)
Birds 59 [30; 60] R 20 x Northward range shift Thomas and Lennon (1999)
Birds 42 [30; 60] R 20 x Absence of range shift Thomas and Lennon (1999)
Lichens 12 [30; 60] R 20 x Increasing richness van Herk et al. (2002)
Lichens 66 [30; 60] R 20 x Decreasing richness van Herk et al. (2002)
Plants 1 [30; 60] R 50 x Northward range shift Walther et al. (2005a, b)
Butterflies 46 [30; 60] R 30 x Decreasing abundance Warren et al. (2001)
Birds 41 [30; 60] R 20 x Absence of range shift Zuckerberg et al. (2009)
Birds 129 [30; 60] R 20 x Northward range shift Zuckerberg et al. (2009)
Birds 43 [30; 60] R 20 x Northward range shift Zuckerberg et al. (2009)
aC: continental; L: local; R: regional.
Examples of recent studies reporting, for both marine and terrestrial ecosystems, modern latitudinal range shifts of organisms at the trailing/rear edge (T/R), the core (C), or the
leading/front edge (L/F) of their latitudinal range. We restricted our literature survey to studies that have explicitly used historical occurrence data to assess range shifts under
modern climate change. We focused on the following two types of historically based studies (Parmesan, 2006): (1) those that directly assess range shifts by comparing historical
and modern data on species distribution across their entire ranges or sections of ranges; and (2) those that infer range shifts by comparing historical and modern data on species
composition or abundance within local communities. Shaded cells represent reports for marine ecosystems, whereas nonshaded cells represent reports for terrestrial ecosystems.
604 Latitudinal and Elevational Range Shifts under Contemporary Climate Change
Latitudinal and Elevational Range Shifts are Laggingbehind Climate Change
It is clear that contemporary climate change is driving lati-
tudinal and elevational range shifts in species distribution
worldwide (Tables 1 and 2). However, the magnitude of these
biotic responses is unlikely to match the magnitude of con-
temporary climate change due to differences in speed,
magnitude, and direction between the climatic changes and
the biotic responses as well as differences among species. Time
lags between geographical range shifts and past climate
changes have already been reported for plants and animals
(Davis, 1986; Svenning and Skov, 2007; Araujo et al., 2008).
One way to assess the importance of such time lags in the
biotic responses to contemporary climate change is to com-
pare the magnitude of observed latitudinal and elevational
Table 2 Examples of observed elevational range shifts under contemporary climate change
Taxonomic
group
No. taxa Latitudinal
zone (deg.)
Spatial
extentaNo. years T/R C L/F Main changes References
Birds 41 [30; 60] L 30 x Absence of range shift Archaux (2004)
Moths 1 [30; 60] L 30 x Upward range shift Battisti et al. (2005)
Trees 3 [30; 60] L 40 x Increasing abundance Beckage et al. (2008)
Trees 3 [30; 60] L 40 x Decreasing abundance Beckage et al. (2008)
Bryophytes 61 [30; 60] R 125 x x x Upward range shift Bergamini et al. (2009)
Moths 102 [0; 30] L 40 x Upward range shift Chen et al. (2009)
Moths 28 [0; 30] L 40 x Downward range shift Chen et al. (2011b)
Moths 28 [0; 30] L 40 x Upward range shift Chen et al. (2011b)
Moths 109 [0; 30] L 40 x Upward range shift Chen et al. (2011b)
Moths 109 [0; 30] L 40 x Upward range shift Chen et al. (2011b)
Plants 64 [30; 60] R 80 x Downward range shift Crimmins et al. (2011)
Epiphyte 1 [30; 60] R 95 x Upward range shift Dobbertin et al. (2005)
Trees 38 [ÿ 30; 0] L 5 x Upward range shift Feeley et al. (2011)
Butterflies 4 [30; 60] R 35 x Upward range shift Franco et al. (2006)
Plants 125 [30; 60] L 95 x x Upward range shift Frei et al. (2010)
Plants 60 [30; 60] L 95 x Increasing richness Grabherr et al. (1994)
Arthropods 280 [30; 60] R 25 x Upward range shift Hickling et al. (2006)
Herptiles 3 [30; 60] R 25 x Downward range shift Hickling et al. (2006)
Fishes 15 [30; 60] R 25 x Upward range shift Hickling et al. (2006)
Birds 22 [30; 60] R 20 x Absence of range shift Hickling et al. (2006)
Mammals 9 [30; 60] R 25 x Upward range shift Hickling et al. (2006)
Butterflies 15 [30; 60] R 35 x Upward range shift Hill et al. (2002)
Plants 140 [30; 60] L 120 x Upward range shift Holzinger et al. (2008)
Plants 22 [0; 30] L 50 x Increasing richness Juvik et al. (2011)
Plants 10 [30; 60] L 30 x x x Upward range shift Kelly and Goulden (2008)
Butterflies 119 [30; 60] R 50 x Upward range shift Konvicka et al. (2003)
Trees 7 [60; 90] L 60 x Upward range shift Kullman (2002)
Plants 171 [30; 60] R 100 x Upward range shift Lenoir et al. (2008)
Plants 60 [30; 60] R 20 x x x Upward range shift Lenoir et al. (2010b)
Plants 22 [ÿ 90; –60] L 40 x x Upward range shift le Roux and McGeoch (2008)
Mammals 28 [30; 60] L 90 x x x Upward range shift Moritz et al. (2008)
Plants 78 [60; 90] R 40 x Upward range shift Odland et al. (2010)
Plants 93 [30; 60] L 50 x Upward range shift Parolo and Rossi (2008)
Plants 10 [30; 60] L 10 x Increasing abundance Pauli et al. (2007)
Plants 10 [30; 60] L 10 x Decreasing abundance Pauli et al. (2007)
Birds 94 [0; 30] 30 x Upward range shift Peh (2007)
Birds 94 [0; 30] 30 x Absence of range shift (Peh 2007)
Amphibians 50 [0; 30] L 5 x x x Decreasing abundance Pounds et al. (1997)
Reptiles 30 [ÿ 30; 0] R 10 x x x Upward range shift Raxworthy et al. (2008)
Birds 1 [30; 60] R 30 x x Upward range shift Tryjanowski et al. (2005)
Plants 31 [30; 60] L 100 x Increasing richness Vittoz et al. (2008)
Plants 18 [30; 60] L 100 x Increasing richness Walther et al. (2005a)
Butterflies 16 [30; 60] R 35 x Upward range shift Wilson et al. (2005)
Birds 41 [30; 60] R 20 x Absence of range shift Zuckerberg et al. (2009)
Birds 129 [30; 60] R 20 x Downward range shift Zuckerberg et al. (2009)
Birds 43 [30; 60] R 20 x Absence of range shift Zuckerberg et al. (2009)
aC: continental; L: local; R: regional.
Examples of recent studies reporting, for terrestrial ecosystems, modern elevational range shifts of organisms at the trailing/rear edge (T/R), the core (C), or the leading/front edge
(L/F) of their elevational range. We restricted our literature survey to studies that have explicitly used historical occurrence data to assess range shifts under modern climate change
(see Table 1 for further explanations).
Latitudinal and Elevational Range Shifts under Contemporary Climate Change 605
Impacts and Distribution of. Migration. Phenological Shifts in
Animals Under Contemporary Climate Change. Species Distribution
Modeling. Translocation as a conservation strategy. Trophic Cascades
References
Ackerly DD, Loarie SR, Cornwell WK, et al. (2010) The geography of climatechange: Implications for conservation biogeography. Diversity and Distributions16: 476–487.
Angert AL, Crozier LG, Rissler LJ, Gilman SE, Tewksbury JJ, and Chunco AJ(2011) Do species’ traits predict recent shifts at expanding range edges?Ecology Letters 14: 677–689.
Araujo MB and Luoto M (2007) The importance of biotic interactions for modellingspecies distributions under climate change. Global Ecology and Biogeography16: 743–753.
Araujo MB, Nogues-Bravo D, Diniz-Filho JAF, Haywood AM, Valdes PJ, andRahbek C (2008) Quaternary climate changes explain diversity among reptilesand amphibians. Ecography 31: 8–15.
Archaux F (2004) Breeding upwards when climate is becoming warmer: no birdresponse in the French Alps. Ibis 146: 138–144.
Atkinson A, Siegel V, Pakhomov E, and Rothery P (2004) Long-term decline in krillstock and increase in salps within the Southern Ocean. Nature 432: 100–103.
Battisti A, Stastny M, Netherer S, et al. (2005) Expansion of geographic range inthe pine processionary moth caused by increased winter temperatures.Ecological Applications 15: 2084–2096.
Beare D, Burns F, Jones E, et al. (2004) An increase in the abundance of anchoviesand sardines in the north-western North Sea since 1995. Global Change Biology10: 1209–1213.
Beaugrand G, Brander KM, Lindley JA, Souissi S, and Reid PC (2003) Planktoneffect on cod recruitment in the North Sea. Nature 426: 661–664.
Beaugrand G, Reid PC, Ibanez F, Lindley JA, and Edwards M (2002) Reorganizationof North Atlantic marine copepod biodiversity and climate. Science 296:1692–1694.
Beckage B, Osborne B, Gavin DG, Pucko C, Siccama T, and Perkins T (2008) Arapid upward shift of a forest ecotone during 40 years of warming in the GreenMountains of Vermont. Proceedings of the National Academy of Sciences of theUnited States of America 105: 4197–4202.
Bergamini A, Ungricht S, and Hofmann H (2009) An elevational shift of cryophilousbryophytes in the last century – an effect of climate warming? Diversity andDistributions 15: 871–879.
Blach-Overgaard A, Svenning JC, Dransfield J, Greve M, and Balslev H (2010)Determinants of palm species distributions across Africa: The relative roles ofclimate, nonclimatic environmental factors, and spatial constraints. Ecography33: 380–391.
Breshears DD, Huxman TE, Adams HD, Zou CB, and Davison JE (2008) Vegetationsynchronously leans upslope as climate warms. Proceedings of the NationalAcademy of Sciences of the United States of America 105: 11591–11592.
Brommer JE (2004) The range margins of northern birds shift poleward. AnnalesZoologici Fennici 41: 391–397.
Brown JH and Lomolino MV (1998) Biogeography. Sunderland: Sinauer Associates.Cardenas ML, Gosling WD, Sherlock SC, Poole I, Pennington RT, and Mothes P
(2011) The response of vegetation on the Andean flank in western Amazonia toPleistocene climate change. Science 331: 1055–1058.
Case TJ, Holt RD, McPeek MA, and Keitt TH (2005) The community context ofspecies’ borders: Ecological and evolutionary perspectives. Oikos 108: 28–46.
Chen IC, Hill JK, Ohlemuller R, Roy DB, and Thomas CD (2011a) Rapid rangeshifts of species associated with high levels of climate warming. Science 333:1024–1026.
Chen IC, Hill JK, Shiu HJ, et al. (2011b) Asymmetric boundary shifts of tropicalmontane Lepidoptera over four decades of climate warming. Global Ecology andBiogeography 20: 34–45.
Chen IC, Shiu HJ, Benedick S, et al. (2009) Elevation increases in mothassemblages over 42 years on a tropical mountain. Proceedings of the NationalAcademy of Sciences of the United States of America 106: 1479–1483.
Colwell RK, Brehm G, Cardelus CL, Gilman AC, and Longina JT (2008) Globalwarming, elevational range shifts, and lowland biotic attrition in the wet tropics.Science 322: 258–261.
Colwell RK and Rangel TF (2009) Hutchinson’s duality: The once and future niche.
Proceedings of the National Academy of Sciences of the United States of
America 106: 19651–19658.Crimmins SM, Dobrowski SZ, Greenberg JA, Abatzoglou JT, and Mynsberge AR
(2011) Changes in climatic water balance drive downhill shifts in plant species’
optimum elevations. Science 331: 324–327.Davis MB (1986) Climatic instability, time lags, and community disequilibrium. In:
Diamond J and Case TJ (eds.) Community Ecology, pp. 269–284. New York:
Harper and Row.Davis MB and Shaw RG (2001) Range shifts and adaptive responses to Quaternary
climate change. Science 292: 673–679.Deutsch CA, Tewksbury JJ, Huey RB, et al. (2008) Impacts of climate warming on
terrestrial ectotherms across latitude. Proceedings of the National Academy of
Sciences of the United States of America 105: 6668–6672.Dobbertin M, Hilker N, Rebetez M, Zimmermann NE, Wohlgemuth T, and Rigling A
(2005) The upward shift in altitude of pine mistletoe (Viscum album ssp.
austriacum) in Switzerland – the result of climate warming? International Journal
of Biometeorology 50: 40–47.Engler R, Randin CF, Vittoz P, et al. (2009) Predicting future distributions of
mountain plants under climate change: Does dispersal capacity matter?
Ecography 32: 34–45.Feeley KJ, Silman MR, Bush MB, et al. (2011) Upslope migration of Andean trees.
Journal of Biogeography 38: 783–791.Forero-Medina G, Joppa L, and Pimm SL (2011) Constraints to species’ elevational
range shifts as climate changes. Conservation Biology 25: 163–171.Franco AMA, Hill JK, Kitschke C, et al. (2006) Impacts of climate warming and
habitat loss on extinctions at species’ low-latitude range boundaries. Global
Change Biology 12: 1545–1553.Frei E, Bodin J, and Walther GR (2010) Plant species’ range shifts in mountainous
areas-all uphill from here? Botanica Helvetica 120: 117–128.Grabherr G, Gottfried M, and Pauli H (1994) Climate effects on mountain plants.
Nature 369: 448.Graham RW (1986) Response of mammalian communities to environmental changes
during the late Quaternary. In: Diamond JM and Case TJ (eds.) Community
Ecology, pp. 300–313. New York: Harper and Row.Hampe A and Petit RJ (2005) Conserving biodiversity under climate change: The
rear edge matters. Ecology Letters 8: 461–467.Heller NE and Zavaleta ES (2009) Biodiversity management in the face of climate
change: A review of 22 years of recommendations. Biological Conservation 142:
14–32.van Herk CM, Aptroot A, and van Dobben HF (2002) Long-term monitoring in the
Netherlands suggests that lichens respond to global warming. Lichenologist 34:
141–154.Hickling R, Roy DB, Hill JK, Fox R, and Thomas CD (2006) The distributions of a
wide range of taxonomic groups are expanding poleward. Global Change
Biology 12: 450–455.Hickling R, Roy DB, Hill JK, and Thomas CD (2005) A northward shift of range
margins in British Odonata. Global Change Biology 11: 502–506.Hill JK, Thomas CD, Fox R, et al. (2002) Responses of butterflies to twentieth
century climate warming: Implications for future ranges. Proceedings of the
Royal Society of London. Series B, Biological Sciences 269: 2163–2171.Hitch AT and Leberg PL (2007) Breeding distributions of North American bird
species moving north as a result of climate change. Conservation Biology 21:
534–539.Hoegh-Guldberg O, Hughes L, McIntyre S, et al. (2008) Assisted colonization and
rapid climate change. Science 321: 345–346.Holbrook SJ, Schmitt RJ, and Stephens JS (1997) Changes in an assemblage of
temperate reef fishes associated with a climate shift. Ecological Applications 7:
1299–1310.Holzinger B, Hulber K, Camenisch M, and Grabherr G (2008) Changes in plant
species richness over the last century in the eastern Swiss Alps: elevational
gradient, bedrock effects and migration rates. Plant Ecology 195: 179–196.Honnay O, Verheyen K, Butaye J, Jacquemyn H, Bossuyt B, and Hermy M (2002)
Possible effects of habitat fragmentation and climate change on the range of
forest plant species. Ecology Letters 5: 525–530.Hutchinson GE (1957) Concluding remarks. Cold Spring Harbor Symposia on
Cambridge, UK: Cambridge University Press.IPCC (2007b) Climate Change 2007: The Physical Science Basis. Cambridge, UK:
Cambridge University Press.
Latitudinal and Elevational Range Shifts under Contemporary Climate Change 609
Author's personal copy
IUCN (2004) 2004 IUCN Red List of Threatened Species: A Global Species
Assessment. Cambridge and Gland, UK and CH: IUCN Publications Services
Unit and the IUCN Species Programme.Jackson ST and Sax DF (2010) Balancing biodiversity in a changing environment:
Extinction debt, immigration credit and species turnover. Trends in Ecology &Evolution 25: 153–160.
Jump AS, Matyas C, and Penuelas J (2009) The altitude-for-latitude disparity in the
range retractions of woody species. Trends in Ecology & Evolution 24:
694–701.Jurasinski G and Kreyling J (2007) Upward shift of alpine plants increases floristic
similarity of mountain summits. Journal of Vegetation Science 18: 711–718.Juvik JO, Rodomsky BT, Price JP, Hansen EW, and Kueffer C (2011) The upper
limit of vegetation on Mauna Loa, Hawaı: A 50th-anniversary reassessment.
Ecology 92: 518–525.Keith DA, Akcakaya HR, Thuiller W, et al. (2008) Predicting extinction risks under
climate change: Coupling stochastic population models with dynamic bioclimatic
habitat models. Biology Letters 4: 560–563.Keith SA, Newton AC, Herbert RJH, Morecroft MD, and Bealey CE (2009) Non-
analogous community formation in response to climate change. Journal for
Nature Conservation 17: 228–235.Kelly AE and Goulden ML (2008) Rapid shifts in plant distribution with recent
climate change. Proceedings of the National Academy of Sciences of the United
States of America 105: 11823–11826.Konvicka M, Maradova M, Benes J, Fric Z, and Kepka P (2003) Uphill shifts in
distribution of butterflies in the Czech Republic: Effects of changing climate
detected on a regional scale. Global Ecology and Biogeography 12: 403–410.Krajick K (2004) Climate change: All downhill from here? Science 303: 1600–1602.Kullman L (2002) Rapid recent range-margin rise of tree and shrub species in the
Swedish Scandes. Journal of Ecology 90: 68–77.Kunstler G, Thuiller W, Curt T, et al. (2007) Fagus sylvatica L. recruitment across a
fragmented Mediterranean Landscape, importance of long distance effective
dispersal, abiotic conditions, and biotic interactions. Diversity and Distributions
13: 799–807.La Sorte FA and Thompson FR (2007) Poleward shifts in winter ranges of North
American birds. Ecology 88: 1803–1812.Lenoir J, Gegout JC, Dupouey JL, Bert D, and Svenning JC (2010b) Forest plant
community changes during 1989–2007 in response to climate warming in the
Jura Mountains (France and Switzerland). Journal of Vegetation Science 21:
949–964.Lenoir J, Gegout JC, Guisan A, et al. (2010a) Going against the flow: Potential
mechanisms for unexpected downslope range shifts in a warming climate.
Ecography 33: 295–303.Lenoir J, Gegout JC, Marquet PA, de Ruffray P, and Brisse H (2008) A significant
upward shift in plant species optimum elevation during the twentieth century.
Science 320: 1768–1771.Lesica P and McCune B (2004) Decline of arctic-alpine plants at the southern
margin of their range following a decade of climatic warming. Journal of
Vegetation Science 15: 679–690.Lima FP, Ribeiro PA, Queiroz N, Hawkins SJ, and Santos AM (2007) Do
distributional shifts of northern and southern species of algae match the
warming pattern? Global Change Biology 13: 2592–2604.Lindgren E, Talleklint L, and Polfeldt T (2000) Impact of climatic change on
the northern latitude limit and population density of the disease-transmitting
European tick Ixodes ricinus. Environmental Health Perspectives 108:
119–123.Loarie SR, Duffy PB, Hamilton H, Asner GP, Field CB, and Ackerly DD (2009) The
velocity of climate change. Nature 462: 1052–1111.Maggini R, Lehmann A, Kery M, et al. (2011) Are Swiss birds tracking climate
change? Detecting elevational shifts using response curve shapes. Ecological
Modelling 222: 21–32.Morin X and Lechowicz MJ (2008) Contemporary perspectives on the niche that
can improve models of species range shifts under climate change. Biology
Letters 4: 573–576.Moritz C, Patton JL, Conroy CJ, Parra JL, White GC, and Beissinger SR (2008)
Impact of a century of climate change on small-mammal communities in
Yosemite National Park, USA. Science 322: 261–264.Morueta-Holme N, Flojgaard C, and Svenning JC (2011) Climate change risks and
conservation implications for a threatened small-range mammal species. Plos
ONE 5: e10360.Mueller RC, Scudder CM, Porter ME, Trotter RT, Gehring CA, and Whitham TG
(2005) Differential tree mortality in response to severe drought: Evidence for
long-term vegetation shifts. Journal of Ecology 93: 1085–1093.
Murphy HT, VanDerWal J, and Lovett-Doust J (2006) Distribution of abundance
across the range in eastern North American trees. Global Ecology and
Biogeography 15: 63–71.Myers P, Lundrigan BL, Hoffman SMG, Haraminac AP, and Seto SH (2009)
Climate-induced changes in the small mammal communities of the Northern
Great Lakes Region. Global Change Biology 15: 1434–1454.Odland A, Hoitomt T, and Olsen SL (2010) Increasing Vascular Plant Richness on
13 High Mountain Summits in Southern Norway since the Early 1970s. Arctic
Antarctic and Alpine Research 42: 458–470.Parmesan C (2006) Ecological and evolutionary responses to recent climate change.
Annual Review of Ecology Evolution and Systematics 37: 637–669.Parmesan C, Ryrholm N, Stefanescu C, et al. (1999) Poleward shifts in
geographical ranges of butterfly species associated with regional warming.
Nature 399: 579–583.Parmesan C and Yohe G (2003) A globally coherent fingerprint of climate change
impacts across natural systems. Nature 421: 37–42.Parolo G and Rossi G (2008) Upward migration of vascular plants following
a climate warming trend in the Alps. Basic and Applied Ecology 9:
100–107.Pauli H, Gottfried M, Reiter K, Klettner C, and Grabherr G (2007) Signals of range
expansions and contractions of vascular plants in the high Alps: observations
(1994–2004) at the GLORIA� master site Schrankogel, Tyrol, Austria. Global
Change Biology 13: 147–156.Pearson RG and Dawson TP (2003) Predicting the impacts of climate change on
the distribution of species: Are bioclimate envelope models useful? Global
Ecology & Biogeography 12: 361–371.Peh KSH (2007) Potential effects of climate change on elevational distributions of
tropical birds in Southeast Asia. The Condor 109: 437–441.Perry AL, Low PJ, Ellis JR, and Reynolds JD (2005) Climate change and
distribution shifts in marine fishes. Science 308: 1912–1915.Post E, Forchhammer MC, Bret-Harte MS, et al. (2009) Ecological dynamics
across the Arctic associated with recent climate change. Science 325:
1355–1358.Pounds JA, Fogden MPL, Savage JM, and Gorman GC (1997) Tests of null models
for amphibian declines on a tropical mountain. Conservation Biology 11:
1307–1322.Pulliam HR (2000) On the relationship between niche and distribution. Ecology
Letters 3: 349–361.R Development Core Team, I (I, 2010) R: A Language and Environment for
Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.Raxworthy CJ, Pearson RG, Rabibisoa N, et al. (2008) Extinction vulnerability of
tropical montane endemism from warming and upslope displacement: A
preliminary appraisal for the highest massif in Madagascar. Global Change
Biology 14: 1703–1720.Ricciardi A and Simberloff D (2009) Assisted colonization: Good intentions and
dubious risk assessment. Trends in Ecology & Evolution 24: 476–477.Richardson AJ and Schoeman DS (2004) Climate impact on plankton ecosystems
in the northeast Atlantic. Science 305: 1609–1612.le Roux PC and McGeoch MA (2008) Rapid range expansion and community
reorganization in response to warming. Global Change Biology 14:
2950–2962.Sagarin RD, Barry JP, Gilman SE, and Baxter CH (1999) Climate-related change in
an intertidal community over short and long time scales. Ecological Monographs
69: 465–490.Sala OE, Chapin III FS, Armesto JJ, et al. (2000) Global biodiversity scenarios for
the year 2100. Science 287: 1770–1774.Scherrer D and Korner C (2011) Topographically controlled thermal-habitat
differentiation buffers alpine plant diversity against climate warming. Journal of
Biogeography 38: 406–416.Smith RIL (1994) Vascular plants as bioindicators of regional warming in
Antarctica. Oecologia 99: 322–328.Soberon J and Nakamura M (2009) Niches and distributional areas: Concepts,
methods, and assumptions. Proceedings of the National Academy of Sciences of
the United States of America 106: 19644–19650.Sturm M, Racine C, and Tape K (2001) Climate change – Increasing shrub
abundance in the Arctic. Nature 411: 546–547.Svenning JC (2003) Deterministic Plio-Pleistocene extinctions in the European
cool-temperate tree flora. Ecology Letters 6: 646–653.Svenning JC and Condit R (2008) Biodiversity in a warmer world. Science 322:
206–207.Svenning JC and Skov F (2004) Limited filling of the potential range in European
tree species. Ecology Letters 7: 565–573.
610 Latitudinal and Elevational Range Shifts under Contemporary Climate Change
Author's personal copy
Svenning JC and Skov F (2007) Ice age legacies in the geographical distributionof tree species richness in Europe. Global Ecology & Biogeography 16: 234–245.
Thomas CD (2011) Translocation of species, climate change, and the end of tryingto recreate past ecological communities. Trends in Ecology & Evolution 26:216–221.
Thomas CD and Lennon JJ (1999) Birds extend their ranges northwards. Nature399: 213.
Tingley MW and Beissinger SR (2009) Detecting range shifts from historicalspecies occurrences: New perspectives on old data. Trends in Ecology &Evolution 24: 625–633.
Tryjanowski P, Sparks TH, and Profus P (2005) Uphill shifts in the distribution ofthe white stork Ciconia ciconia in southern Poland: the importance of nestquality. Diversity and Distributions 11: 219–223.
Vittoz P, Bodin J, Ungricht S, Burga CA, and Walther GR (2008) One century ofvegetation change on Isla Persa, a nunatak in the Bernina massif in the SwissAlps. Journal of Vegetation Science 6: 671–680.
Walther GR (2010) Community and ecosystem responses to recent climate change.Philosophical Transactions of the Royal Society B – Biological Sciences 365:2019–2024.
Walther GR, Beibner S, and Burga CA (2005a) Trends in the upward shift of alpineplants. Journal of Vegetation Science 16: 541–548.
Walther GR, Berger S, and Sykes MT (2005b) An ecological ‘footprint’ of climatechange. Proceedings of the Royal Society of London. Series B, BiologicalSciences 272: 1427–1432.
Warren MS, Hill JK, Thomas JA, et al. (2001) Rapid responses of British butterfliesto opposing forces of climate and habitat change. Nature 414: 65–68.
Wilson RJ, Gutierrez D, Gutierrez J, Martinez D, Agudo R, and Monserrat VJ (2005)Changes to the elevational limits and extent of species ranges associated withclimate change. Ecology Letters 8: 1138–1146.
Zuckerberg B, Woods AM, and Porter WF (2009) Poleward shifts in breeding birddistributions in New York State. Global Change Biology 15: 1866–1883.
Latitudinal and Elevational Range Shifts under Contemporary Climate Change 611