Ecological Research (1996) 11,229-247 The effects of a typhoon on Japanese warm temperate rainforests P.J. B ELLINGHAM, ~ TAKASHI KOHYAMA T AND SHIN-ICHIRO AIBA t Center for Ecological Research, Kyoto University, Shimosakamoto, Otsu 5 20-O l , Japan A powerful typhoon (Typhoon No. 13) affected stands of primary warm temperate rainforest in Yakushima, southern Japan, in 1993. Censuses in three sites 1-5 months after the typhoon showed low levels of mortality resulting from the typhoon (0.4-3.0% of stems). Stems killed by the typhoon were generally larger than surviving stems. Among surviving stems there were generally low levels of damage (e.g. 0.5-1.3% of surviving stems lost crowns) and not all damage was widespread (e.g. defoliation was apparent only in one of three sites). The sizes of uprooted stems, stems that lost crowns and relatively undamaged stems were not different. Different species appeared to be damaged in different ways at different sites. Damage was most evident in higher altitude seaward sites but gap formation was more frequent in lower altitude sites near valley floors. After the typhoon the resultant gap area occupied 9.4% of one site and 8.6% of another, which is greater than that before the typhoon. Fresh sprouts were found on 17.35 % of 2161 stems after the typhoon, including many apparently undamaged stems as well as those that were damaged. Species which sprouted most frequently were those that regenerate by seed least frequently in these forests; these species may maintain their relative abundance in part by sprouting. Most tree species in these forests may be relatively resistant to typhoons and there may be more opportunities for their regeneration following gap formation caused by the typhoon. Key words: damage; disturbance; gaps; mortality; sprouting; resistance. INTRODUCTION Disturbance by wind can have important implica- tions for forests (White 1979). Strong winds may alter forest structure and may produce treefall gaps ranging in size from relatively small to very large (e.g. Foster 1988a; Webb 1989). The effects of strong winds on trees may vary considerably among species (e.g. Zimmerman et al. 1994; Bellingham et al. 1995). The mortality of particular tree species may be very high, and substantial changes in forest composition may result (Foster 1988b). After strong winds affect forests, opportunities may arise *Present address: Manaaki Whenua-Landcare Research New Zealand Limited, P.O. Box 69, Lincoln 8152, New Zealand. *Present address: Graduate School of Environmental Earth Science, Hokkaido University, Kita-ku, Sapporo 060, Japan. Received 8 May 1995. Accepted 24 November 1995. for other changes in forest composition, such as colonization by species that regenerate in high light environments (e.g. Whitmore 1974). There have been several recent studies which have drawn attention to the short-term and long-term effects of typhoons, cyclones and hurricanes in tropical and temperate forests (e.g. Whitmore 1974; Shaw 1983; Dittus 1985; Weaver 1986; Foster 1988a,b; Brokaw & Walker 1991; Merrens & Peart 1992; Elmqvist et al. 1994). Cydones are most frequent in the north-west Pacific, where they are known as typhoons. In Japan, the areas most frequently affected by typhoons are the southern part of the main islands and the subtropical southern islands. Several studies have concluded that typhoons are important causes of gaps in some Japanese forests and are, thus, important for the regeneration of several tree species (e.g. Suzuki & Tsukahara 1987; Yamamoto 1992). Naka (1982) reported damage caused by a typhoon in an ever- green, broad-leaved forest in south-western Honshu and believed that treefall gaps created during
Transcript
Ecological Research (1996) 1 1 , 2 2 9 - 2 4 7
The effects of a typhoon on Japanese warm temperate rainforests
P.J. B ELLINGHAM, ~ TAKASHI KOHYAMA T AND SHIN-ICHIRO AIBA t
Center for Ecological Research, Kyoto University, Shimosakamoto, Otsu 5 20-O l , Japan
A powerful typhoon (Typhoon No. 13) affected stands of primary warm temperate rainforest in Yakushima, southern Japan, in 1993. Censuses in three sites 1-5 months after the typhoon showed low levels of mortality resulting from the typhoon (0.4-3.0% of stems). Stems killed by the typhoon were generally larger than surviving stems. Among surviving stems there were generally low levels of damage (e.g. 0.5-1.3% of surviving stems lost crowns) and not all damage was widespread (e.g. defoliation was apparent only in one of three sites). The sizes of uprooted stems, stems that lost crowns and relatively undamaged stems were not different. Different species appeared to be damaged in different ways at different sites. Damage was most evident in higher altitude seaward sites but gap formation was more frequent in lower altitude sites near valley floors. After the typhoon the resultant gap area occupied 9.4% of one site and 8.6% of another, which is greater than that before the typhoon. Fresh sprouts were found on 17.35 % of 2161 stems after the typhoon, including many apparently undamaged stems as well as those that were damaged. Species which sprouted most frequently were those that regenerate by seed least frequently in these forests; these species may maintain their relative abundance in part by sprouting. Most tree species in these forests may be relatively resistant to typhoons and there may be more opportunities for their regeneration following gap formation caused by the typhoon.
Disturbance by wind can have important implica- tions for forests (White 1979). Strong winds may alter forest structure and may produce treefall gaps ranging in size from relatively small to very large (e.g. Foster 1988a; Webb 1989). The effects of strong winds on trees may vary considerably among species (e.g. Zimmerman et al. 1994; Bellingham et al. 1995). The mortality of particular tree species may be very high, and substantial changes in forest composition may result (Foster 1988b). After strong winds affect forests, opportunities may arise
*Present address: Manaaki Whenua-Landcare Research New Zealand Limited, P.O. Box 69, Lincoln 8152, New Zealand.
*Present address: Graduate School of Environmental Earth Science, Hokkaido University, Kita-ku, Sapporo 060, Japan.
Received 8 May 1995. Accepted 24 November 1995.
for other changes in forest composition, such as colonization by species that regenerate in high light environments (e.g. Whitmore 1974).
There have been several recent studies which have drawn attention to the short-term and long-term effects of typhoons, cyclones and hurricanes in tropical and temperate forests (e.g. Whitmore 1974; Shaw 1983; Dittus 1985; Weaver 1986; Foster 1988a,b; Brokaw & Walker 1991; Merrens & Peart 1992; Elmqvist et al. 1994). Cydones are most frequent in the north-west Pacific, where they are known as typhoons. In Japan, the areas m o s t frequently affected by typhoons are the southern part of the main islands and the subtropical southern islands. Several studies have concluded that typhoons are important causes of gaps in some Japanese forests and are, thus, important for the regeneration of several tree species (e.g. Suzuki & Tsukahara 1987; Yamamoto 1992). Naka (1982) reported damage caused by a typhoon in an ever- green, broad-leaved forest in south-western Honshu and believed that treefall gaps created during
typhoons were important for the regeneration of these forests. Shimizu (1984) studied the effects of a typhoon in forests of Chichijima, Ogasawara Islands, and considered the dominant tree Distylium lepidotum Nakai to be resistant to typhoons, as very few were damaged compared to the three other common tree species in the same forests, Pandanus boninensis Warb., Pouteria obovata (R.Br.) Baehni and Schima mertensiana Koidz.
Only a few studies have examined the effects of typhoons on Japanese warm temperate rainforests. After a typhoon affected warm temperate rainforest in southern Kyushu, Taoda (1987) found that Castanopsis cuspidata (Thunb.) Schottky was more severely damaged, mostly by breakage, than other common species in the same forest, and predicted that the consequence would be a succession from forest dominated by C cuspidata to one dominated by Distylium racemosum and Quercus salicina. The passage of a powerful typhoon in 1993 near warm temperate rainforests in Yakushima in southern Japan presented an opportunity to document its immediate effects and consider its long-term impli- cations for forest composition. We studied its effects in primary warm temperate rainforests dominated by D. racemosum (species nomenclature for Yaku- shima forests follows Mitsuta and Nagamasu 1984), in which forest composition, structure and dynamics have been the subject of earlier and ongoing studies (e.g. Kohyama 1986, 1992, 1993). In this study, w e address the following questions. What kinds of damage did the typhoon cause? Were there differences in the ways in which trees of different size and species were affected during the typhoon? Were patterns of damage in these forests related to landscape features or to where particular tree species grew? How variable was the damage caused by this typhoon among different sites, and therefore, how predictable are the effects of future typhoons?
Although this study is primarily concerned with the immediate effects of a typhoon on adult trees, we also studied some of the immediate responses of surviving stems. Vegetative sprouting by epicormic shoots has been shown to be a means of recovery from wind damage m tropical forests (e.g. Putz et al. 1983; Unwin et al. 1988; Yih et al. 1991; Belling- ham et al. 1994; Zimmerman et al. 1994). There- fore, we assessed the extent of sprouting in Yakushima forests and its variation among species.
We also considered what the implications of dam- age by this typhoon may be for forest dynamics in future.
S T U D Y SITES A N D METHODS
Study sites
We examined stands of primary warm temperate rainforest on Yakushima, northern Ryukyu Islands, southern Japan. We examined in detail forests studied by Kohyama (1986) on the western slopes of the Segire River valley (30~176 130~ 450-610 m a.s.1.) and in the Koyohji River valley (30~ 130~ 520-700 m a.s.1.; Fig. 1). Both catchments are located in the south-west of Yakushima, on a uniform base of Miocene granite which forms the entire mountain body of the island.
Typhoon No. 13 of 1993 was the third most powerful typhoon to affect the main islands of Japan since 1945 (Yamamoto et al. 1994). It struck Yakushima on 3 September 1993 and its eye passed, at its nearest, 60 km to the north-north-west of the island (Fig. 1) moving at 35 km h -1, with a pressure at the eye of 925 hPa. At the Yakushima Weather Station at Koseda, 2.5 km east-north-east of our study sites, average wind velocity at the height of the typhoon was 21.6 In s -1, clockwise from south-south-west, with maximum speeds of 55.4 m s -1. Wind speeds over 10 m s -1 were recorded for 8 h at the station. Total rainfall at the station during the typhoon was 212.5 m m over a 24 h period with a maximum hourly rainfall during that period of 64 mm. After passing by Yakushima, Typhoon No. 13 passed through southern and eastern Kyushu, Shikoku, and through the Chugoku District of Honshu before weakening in the Sea of Japan (Japanese Government Meteoro- logical Service, unpubl, data; Yamamoto et al. 1994).
Methods
We recorded the effects of the typhoon on trees ~>2 cm d.b.h, between September 1993 and Febru- ary 1994 (i.e. up to 5 months after the typhoon). We recorded mortality and damage to stems caused by the typhoon (after the methods of Bellingham
Fig. 1. Left: Map of south-west Yakushima, showing the location of three study areas. Contour intervals = 100 m. Right: Location of Yakushima in southern Japan with enlarged area shaded. The track of the eye of Typhoon No. 13 of 1993, moving from south-west to north-east, is shown (adapted from Yamamoto et al. 1994).
1991) including: (i) mortality that we could ascribe to the typhoon, recording dead stems only when bark was still present and decay had not com- menced; (ii) broken stems, for which we recorded stems that had completely died above the breakage point ('crown lost'), those in which the crowns were bent over but remained alive, and those from which a substantial part of the crown had been broken ('partial breakage'); (iii) uprooted stems, including only stems uprooted t>40 ~ from their pre-typhoon position with freshly disturbed soil around their root plates; and (iv) severely defoliated stems, in which we recorded stems from which we estimated more than 50% of leaves had been lost. In analysis of defoliated stems, we did not include the few, relatively rare deciduous species of the forest (e.g. Aleurites cordata, Fagara ailanthoides, Stewartia monadelptaa and Styrax japonica) that would
ordinarily have been without leaves during this survey.
w e surveyed damage caused by the typhoon in permanent sample plots (total area 1.53 ha) in
which all trees >12 cm d.b.h, were permanently tagged and identified (Kohyama 1986). We re- corded damage in plots in the Koyohji River valley (0.91 ha; Fig. 1) in September 1993 and in plots on the lower slopes of the Segire River valley (TK plots, 0.62 ha; Fig. 1) in November 1993. From these plots we recorded data on mortality that could be attributed to the typhoon: that is, damage caused to trees, and the incidence of gap formation. Gaps were defined according to Kohyama (1986) to permit comparisons with studies carried out before the typhoon: that is, gap area was calculated as an ellipse derived from two perpendicular axis measurements made directly under the canopy opening.
232 P.J. Bellingham et al.
Our earliest observations in the Segire River valley suggested some areas of forest were more damaged than others. To investigate damage over a wider area of the forests in the Segire River valley, we estab- lished seven parallel 260 m long transects, 50 m apart, oriented north-south on the western slopes of the valley during January-February 1994. We set up 10 m x 10 m plots (PJB plots; Fig. 1) at 50 m intervals along each transect (i.e. 6 plots per transect). We excluded part of the topmost transect which crossed the dividing ridge of the Segire River valley, thus recording 41 plots in total (0.41 ha) from within a total area of ca 7.7 ha. We recorded the aspect, slope, altitude and physiography (spur or ridge crest, flank, gully) of each plot. To increase the total sample of stems we also set up two contiguous 20 m x 20 m plots between two of the higher altitude transects in an area in which many stems were defoliated (0.08 ha). Within all of these plots we recorded d.b.h, and the species of all stems i>2 cm d.b.h. We recorded the dimensions of gaps formed by the typhoon along the seven transects in the PJB study area, incorporating an area up to 5 m either side of the transect. Excluding 55 m of the highest altitude transect, which crossed the dividing ridge of the Segire River catchment, we recorded gaps within a total survey area of 1.77 ha. Within each gap we recorded the d.b.h, and species of stems that had contributed to the gap. In the PJB plots, in addition to measures of mortality and damage, we also recorded the numbers of fresh epicormic sprouts emerging above and below 2.5 m on each stem. Epicormic sprouting was observed in these forests before the passage of Typhoon No. 13, and we distinguished sprouts with lignified stems present before the typhoon in contrast to fresh sprouts which developed after the typhoon. We refer to the sprouts that existed before Typhoon No. 13 as a 'sprout bank', a term coined by Bellingham et al. (1994) to describe sprouts that existed in a Jamaican montane forest before the passage of a hurricane and which grew rapidly afterwards.
Using data from the PJB plots along transects, we analyzed patterns of mortality and severe damage among living stems (combined totals of stems that lost crowns, were uprooted or were severely defoli- ated) with respect to aspect, slope, altitude and physiography, and interactions between these vari- ables. We reduced the continuous variables to categories and analyzed the data using General
Linear Models in sYsTaT 5 (Wilkinson 1989). We examined percentage mortality and damage per plot (arcsine square-root transformed data) for all species combined and for each of three common, ubiqui- tous species.
We also examined whether the typhoon affected some species more than others as a result of where these species most frequently occurred. To do this, we performed three ordinations using basal area data for each species in each of the PJB plots along transects. In the first ordination, we used the total basal area. In the second, we performed the same ordination but subtracted the basal area killed by the typhoon, and in the third ordination, we subtracted from the total basal area both the dead basal area and the basal area of living stems that were severely damaged. The three ordinations were carried out using Canonical Correspondence Analysis (ter Braak 1986), including the four environmental variables collected from each plot. We carried out these analyses using the computer package caNoco (ter Braak 1988).
Values for individual species were compared with the overall value for all species using G-tests (em- ploying Williams' correction where appropriate). Some of the 'significant' differences, especially those at 5%, are a result of random variation so we attach greatest importance to individual comparisons sig- nificant at greater than the 1% level. Other tests of trends among species were made using unpaired t-tests, ANOVA and correlations. Comparisons were made between individual species after aNOVa using Scheff~ F-tests.
RESULTS
Morta l i ty
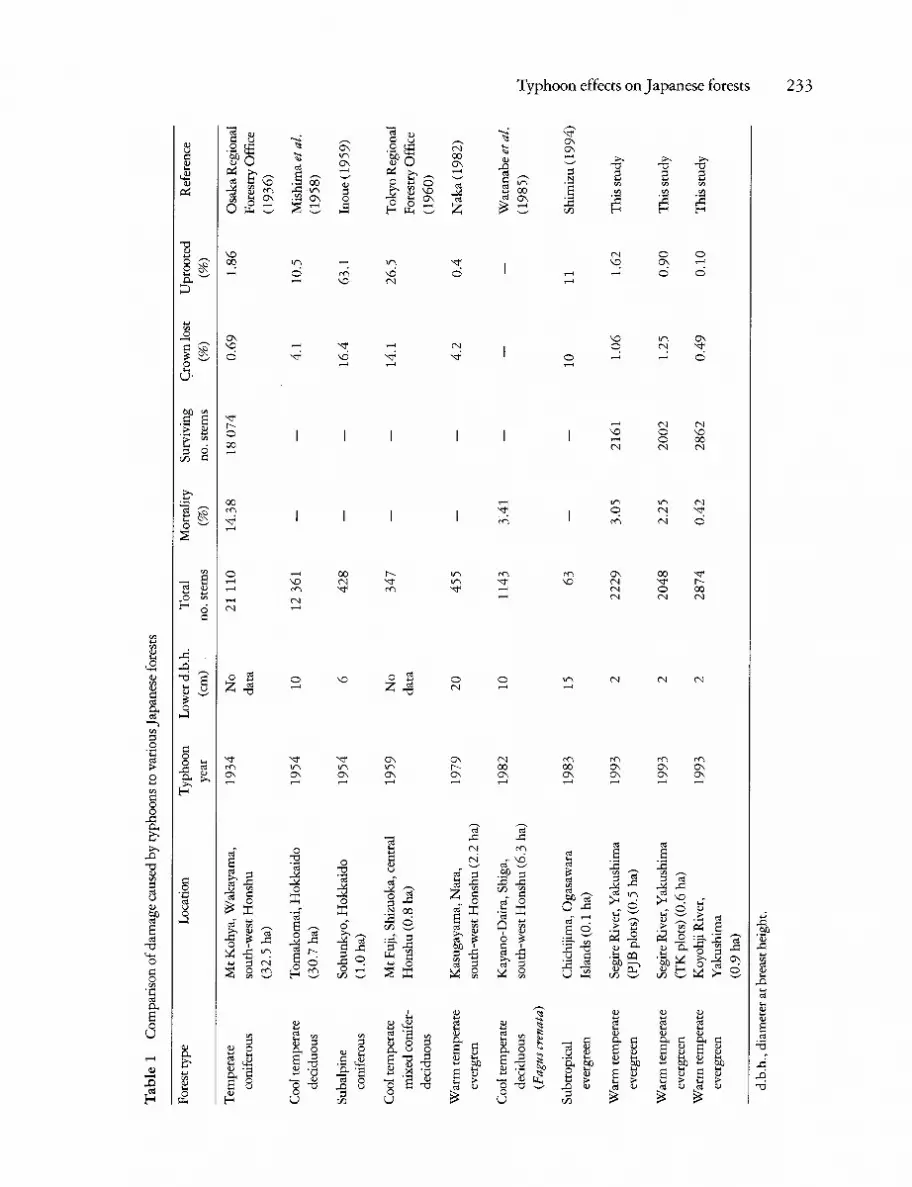
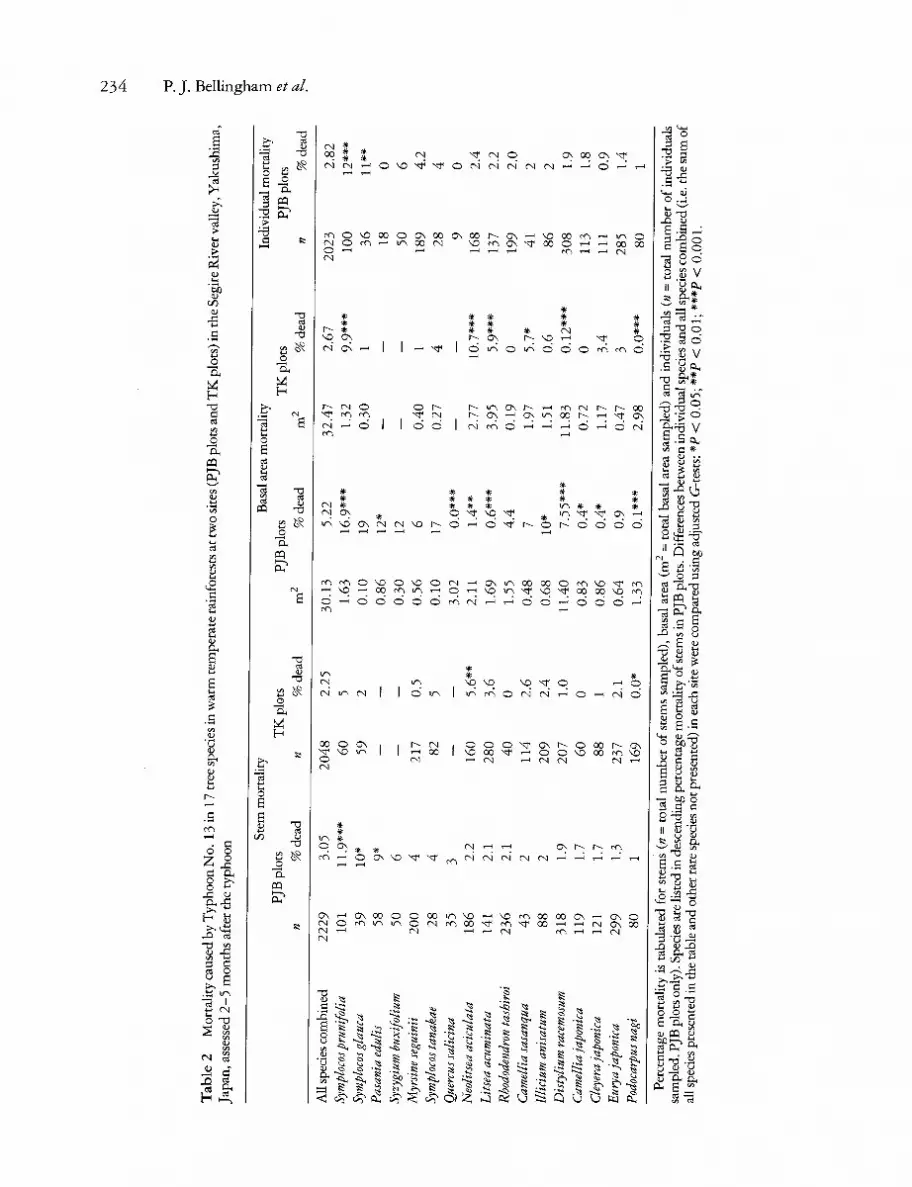
Mortality caused by the typhoon was higher in both sets of plots in the Segire River valley than in the Koyohji River valley (Table 1). In the Segire River valley, levels of mortality were higher in the wide- spread survey (PJB plots) than in the plots near the valley bottom (TK plots). In the PJB plots, the percentage mortality of the total basal area of all species combined exceeded that of stems and indi- viduals (Table 2). In the inventory period that included Typhoon No. 13, annual mortality in the TK plots derived according to the exponential
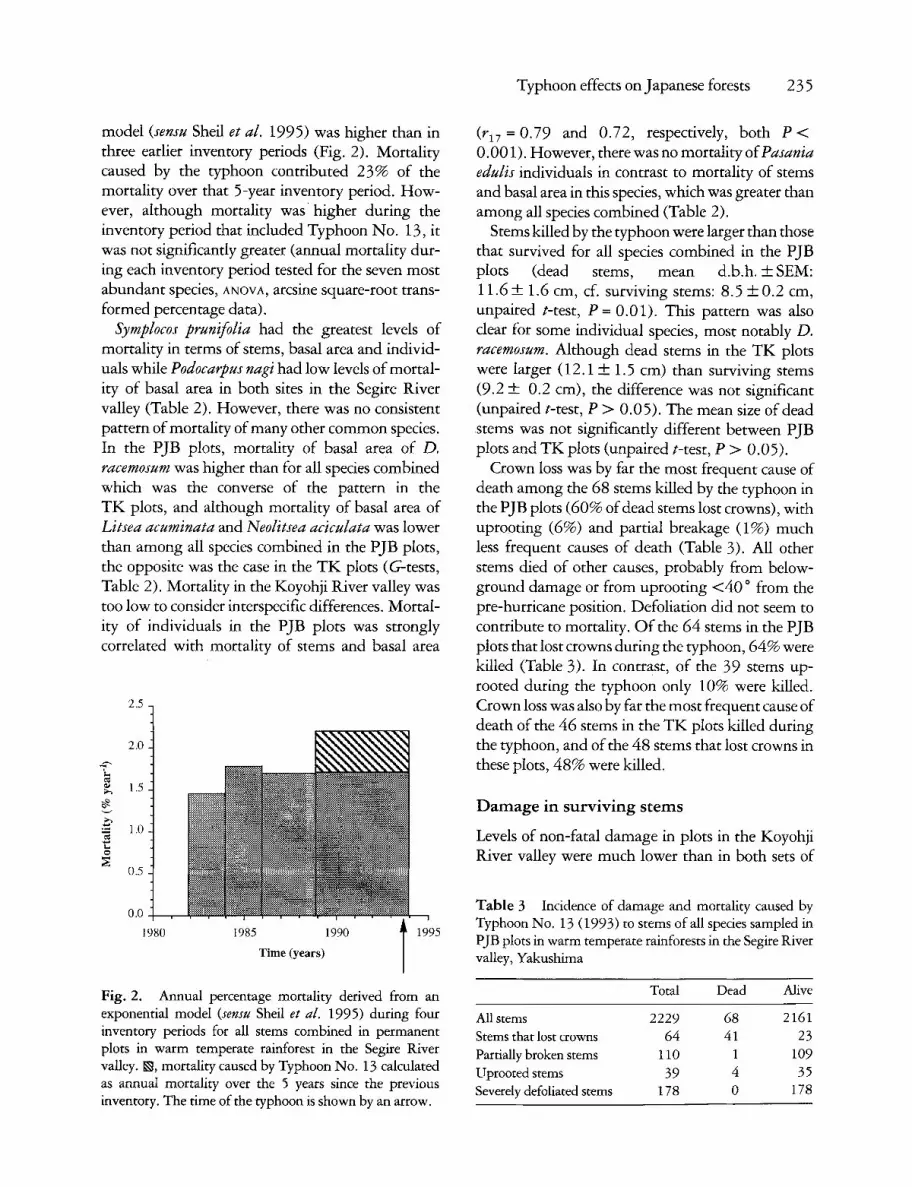
model (sensu Shell et al. 1995) was higher than in three earlier inventory periods (Fig. 2). Mortality caused by the typhoon contributed 23% of the mortality over that 5-year inventory period. How- ever, although mortality was higher during the inventory period that included Typhoon No. 13, it was not significantly greater (annual mortality dur- ing each inventory period tested for the seven most abundant species, ANOVA, arcsine square-root trans- formed percentage data).
Symplocos prunifolia had the greatest levels of mortality in terms of stems, basal area and individ- uals while Podocarpus nagi had low levels of mortal- ity of basal area in both sites in the Segire River valley (Table 2). However, there was no consistent pattern of mortality of many other common species. In the PJB plots, mortality of basal area of D. racemosum was higher than for all species combined which was the converse of the pattern in the TK plots, and although mortality of basal area of Litsea acuminata and Neolitsea aciculata was lower than among all species combined in the PJB plots, the opposite was the case in the TK plots (G-tests, Table 2). Mortality in the Koyohji River valley was too low to consider interspecific differences. Mortal- ity of individuals in the PJB plots was strongly correlated with mortality of stems and basal area
2.5
2.0
1.5
1.0
0.5
0 . 0 i , , , r i
1980 1985 1990
Time (years)
i
1995
Fig. 2. Annual percentage mortality derived from an exponential model (sensu Shell et al. 1995) during four inventory periods for all stems combined in permanent plots in warm temperate rainforest in the Segire River valley. ~, mortality caused by Typhoon No. 13 calculated as annual mortality over the 5 years since the previous inventory. The time of the typhoon is shown by an arrow.
Typhoon effects on Japanese forests 235
(r17 =0 .79 and 0.72, respectively, both P < 0.001). However, there was no mortality of Pasania edulis individuals in contrast to mortality of stems and basal area in this species, which was greater than among all species combined (Table 2).
Stems killed by the typhoon were larger than those that survived for all species combined in the PJB plots (dead stems, mean d.b.h. + SEM: 11.6 + 1.6 cm, cf. surviving stems: 8.5 + 0.2 cm, unpaired t-test, P = 0.01). This pattern was also clear for some individual species, most notably D. racemosum. Although dead stems in the TK plots were larger (12.1 + 1.5 cm) than surviving stems (9.2 + 0.2 cm), the difference was not significant (unpaired t-test, P > 0.05). The mean size of dead stems was not significantly different between PJB plots and TK plots (unpaired t-test, P > 0.05).
Crown loss was by far the most frequent cause of death among the 68 stems killed by the typhoon in the PJB plots (60% of dead stems lost crowns), with uprooting (6%) and partial breakage (1%) much less frequent causes of death (Table 3). All other stems died of other causes, probably from below- ground damage or from uprooting < 4 0 o from the pre-hurricane position. Defoliation did not seem to contribute to mortality. O f the 64 stems in the PJB plots that lost crowns during the typhoon, 64% were killed (Table 3). In contrast , of the 39 stems up- rooted during the typhoon only 10% were killed. Crown loss was also by far the most frequent cause of death of the 46 stems in the TK plots killed during the typhoon, and of the 48 stems that lost crowns in these plots, 48% were killed.
Damage in surviving stems
Levels of non-fatal damage in plots in the Koyohji River valley were much lower than in both sets of
Table 3 Incidence of damage and mortality caused by Typhoon No. 13 (1993) to stems of all species sampled in PJB plots in warm temperate rainforests in the Segire River valley, Yakushima
Total Dead Alive
All stems 2229 68 2161 Stems that lost crowns 64 41 23 Partially broken stems 110 1 109 Uprooted stems 39 4 35 Severely defoliated stems 178 0 178
lxO
vO
o
Ta
ble
4
Fre
quen
cy o
f ty
pes
of n
on-f
atal
seve
re d
amag
e ca
used
by
Ty
ph
oo
n N
o. 1
3 in
sur
vivi
ng st
ems
of
17 t
ree
spec
ies
in w
arm
tem
pera
te r
ainf
ores
ts in
the
Segi
re R
iver
va
lley
, Yak
ushi
ma,
Jap
an,
asse
ssed
2-5
mo
nth
s af
ter
the
typh
oon.
Th
e fr
eque
ncy
of s
prou
ting
in th
ese
spec
ies,
ass
esse
d co
ncur
rent
ly, i
s al
so t
abul
ated
0~
St
ems
that
lost
cro
wns
U
proo
ted
stem
s
PJB
plo
ts
TK
plo
ts
PJB
plo
ts
TK
plo
ts
n al
ive
%
n al
ive
%
n al
ive
%
n al
ive
%
Seve
rely
def
olia
ted
stem
s S
prou
ting
stem
s
PJB
plo
ts
PJB
plo
ts
n al
ive
%
n al
ive
%
All
spec
ies c
ombi
ned
2161
1.
06
2002
1.
25
2161
1.
62
2002
0.
90
2129
8.
36
2161
14
.30
Cam
ellia
japo
nica
11
7 1.
7 60
2
117
1.7
60
2 11
5 7.
0 11
7 17
.9
Cam
ellia
sasa
nqua
42
0
111
2.7
42
0 11
1 1.
8 42
0*
42
7
Cle
yera
japo
nica
11
9 0.
0 87
0
119
1.7
87
0 11
9 10
.1
119
11.8
D
isty
lium
rac
emos
um
312
0.3
205
1.0
312
1.3
205
0.5
311
2.3*
**
312
11.2
E
urya
japo
nica
29
5 0.
7 23
2 1.
3 29
5 1.
4 23
2 2.
6*
293
3.1"
**
295
7.5*
**
Illic
ium
ani
satu
m
86
5**
204
1.0
86
0 20
4 2.
0 82
10
86
17
Lits
eaac
umin
ata
138
0.7
270
1.1
138
0.7
270
0.4
137
5.8
138
22.5
**
Myr
sine
segu
inii
192
0.0
216
1.4
192
1.0
216
0.0
192
4.2*
19
2 6.
3***
N
eolit
seaa
cicu
lata
18
2 2.
2 15
1 2.
6 18
2 2.
2 15
1 0.
0 17
8 13
.5"
182
17.6
P
asan
ia ed
ulis
53
6"
--
--
53
4
--
--
50
16
53
28**
Po
doca
rpus
nagi
79
0
169
1.2
79
3 16
9 0.
6 79
0"
**
79
4**
Que
rcus
salic
ina
34
0 -
- 34
0
- -
34
44**
* 34
53
***
Rhod
oden
dron
tash
iroi
23
1 0.
0 40
0
231
3.5*
40
0
231
17.3
"**
231
3.9*
**
Sym
ploc
osgl
auca
35
0
58
2 35
0
58
0 35
20
**
35
51"*
* Sy
mpl
ocos
prun
ifolia
89
1
57
0 89
1
57
0 88
16
"*
89
18
Sym
ploc
os ta
naka
e 27
0
78
1 27
0
78
3 27
0
27
11
Syzy
gium
bux
ifoliu
m
47
2 --
--
47
4
--
--
46
7 47
43
***
Freq
uenc
y of
cro
wn
loss
and
upr
ooti
ng w
as a
sses
sed
in tw
o si
tes
(PJB
and
TK
plo
ts);
sev
ere
defo
liat
ion
and
spro
utin
g fr
eque
ncy
was
ass
esse
d in
PJB
plo
ts o
nly.
n =
nu
mb
er o
f su
rviv
ing s
tem
s (f
or s
ever
ely
defo
liate
d st
ems,
sur
vivi
ng st
ems
that
lost
crow
ns w
ere
excl
uded
and
am
ong
all s
peci
es co
mbi
ned
deci
duou
s spe
cies
that
wou
ld n
ot u
sual
ly h
ave
had
leav
es
at th
e ti
me
of s
urve
y w
ere
also
exc
lude
d).
Dif
fere
nces
bet
wee
n in
divi
dual
spec
ies a
nd a
ll sp
ecie
s com
bine
d (i
.e. t
he s
um o
f all
spec
ies p
rese
nted
in
the
tabl
e an
d ot
her
rare
spe
cies
not
pr
esen
ted)
in
each
site
wer
e co
mpa
red
usin
g ad
just
ed G
-res
ts: *
P <
0.0
5; *
*P <
0.
01;
***P
< 0
.001
.
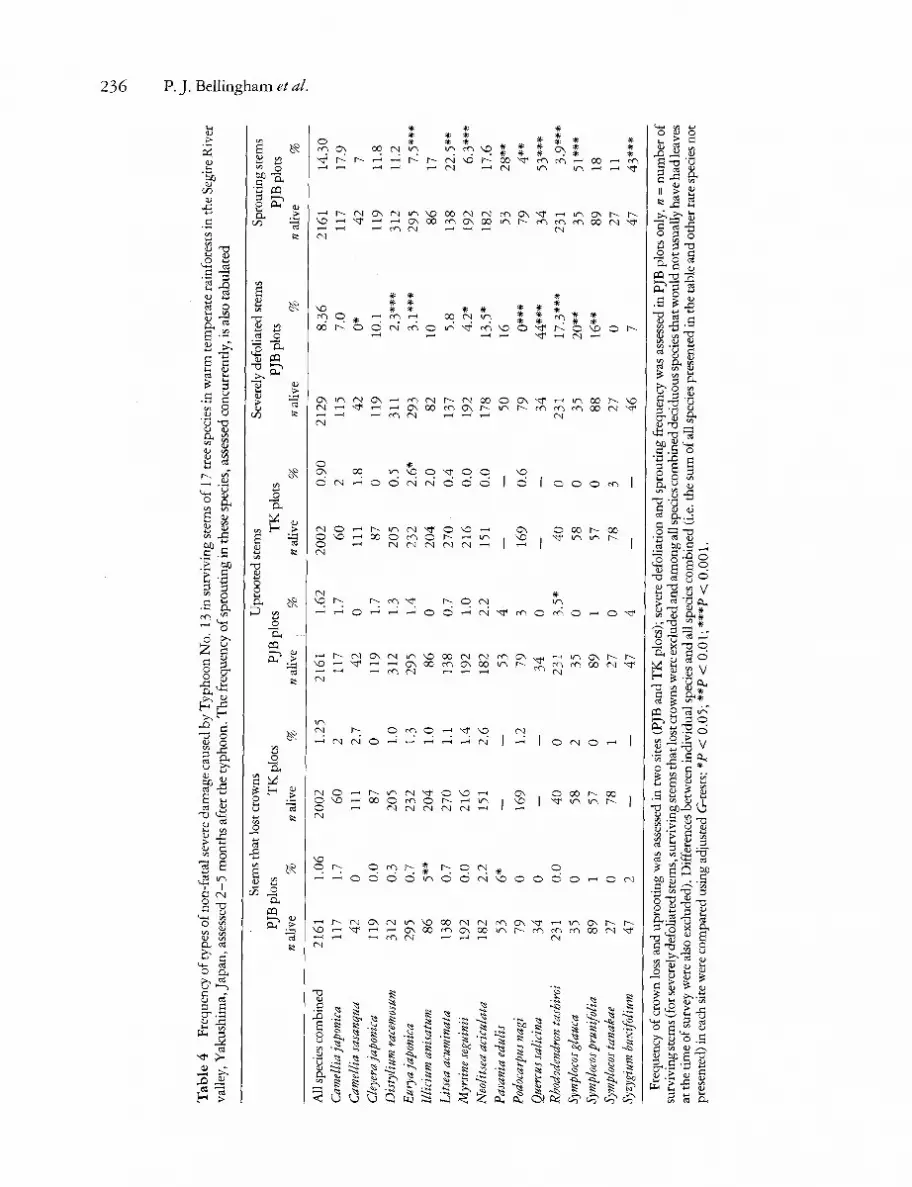
plots in the Segire River valley (Table 1). In the Segire River valley, although crown loss in surviving stems was slightly higher in the T K plots than in the PJB plots (Table 4), other types of damage such as uprooting occurred less frequently. Importantly, in contrast to PJB plots where 8.27% of surviving stems were severely defoliated during the typhoon, no stems in the T K plots were severely defoliated. In the PJB plots, the frequencies with which stems of a given species lost crowns, were uprooted or were severely defoliated were not correlated (tested among 17 common species, P > 0.05). The fre- quency with which stems of particular species were severely defoliated during the typhoon was nega- tively correlated with leaf lamina thickness of the same species (rl l = - 0 . 6 7 , P < 0.05), but was not related to leaf size (data from Grubb et al.
1975). There was no correlation between the two survey
areas in the Segire River valley (PJB plots vs T K plots) in the frequency with which species were killed or lost crowns during the typhoon. Further- more, species frequently uprooted in one survey area were infrequently uprooted in the other and vice versa (r14 = - 0.52, P < 0.05). W e tested whether there were similar levels of damage to species in the T K plots and a subsample of PJB plots near the valley floor at comparable altitude (500-530 m), but neither mortality nor damage among species were correlated between the two sites (P > 0.05). Heights of undamaged stems were surveyed in some of the T K plots after the typhoon, but there was no correlation between mean stem heights of these
Typhoon effects on Japanese forests 23 7
surviving stems and the frequency of mortality, crown loss or uprooting among 14 common species in these plots (P > 0.05).
For all species combined in the PJB plots, partially broken stems and severely defoliated stems were significantly larger than relatively undamaged stems, but there were no differences between the mean sizes of stems that lost crowns, uprooted stems and undamaged stems (Table 5). Similar patterns were found for two individual species in PJB plots D. racemosum and Rhododendron tashiroi. In con- trast, in surviving stems in the T K plots there were no differences in the size of stems that lost crowns, were partially broken, were uprooted, and relatively undamaged stems (Table 5). Surviving stems that lost crowns or were uprooted during the typhoon were not of different sizes in the two survey areas (unpaired t-tests, P > 0.05).
P a t t e r n s o f m o r t a l i t y a n d d a m a g e
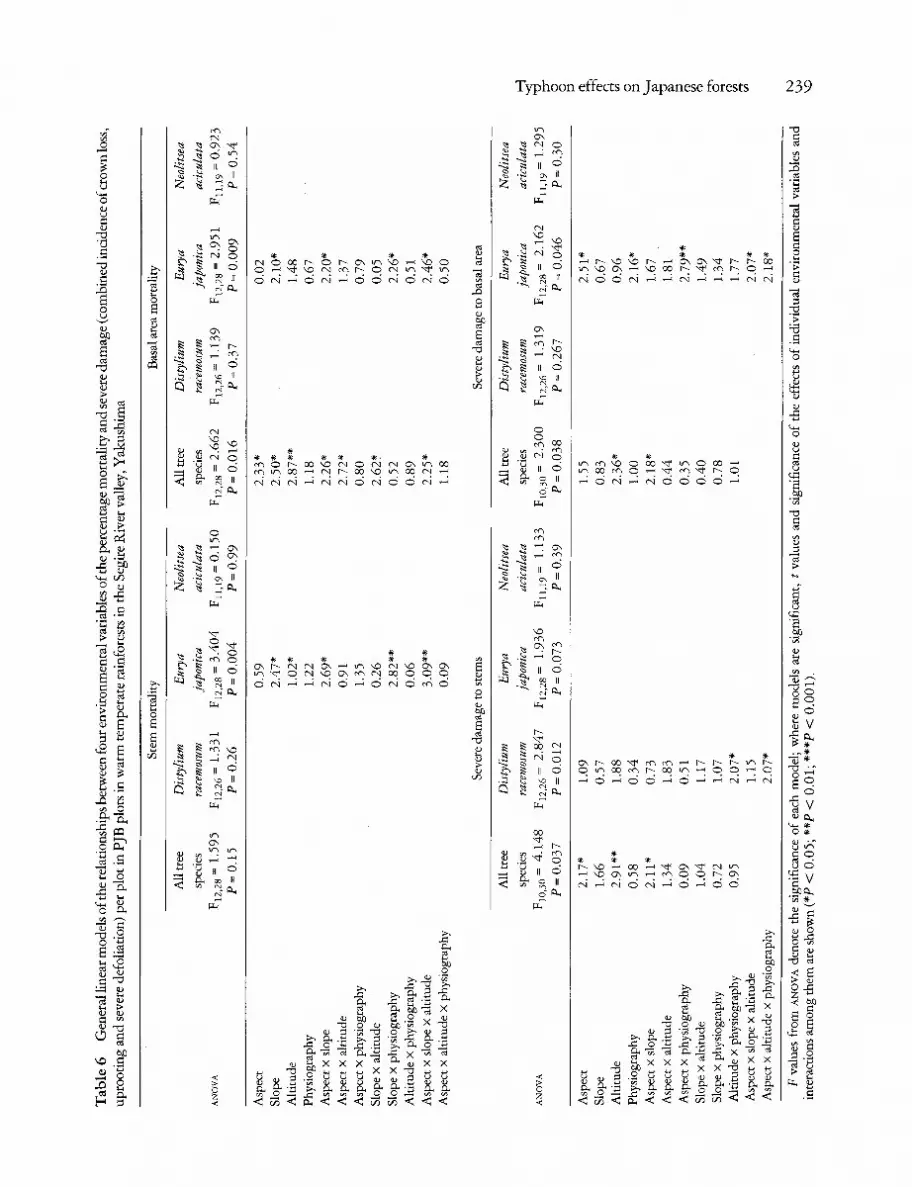
In the Segire River valley, there was greatest mortal- ity of basal area of all species combined in PJB plots at higher altitude ( > 5 0 0 m), facing south-east to south-west and of low to moderate (<35 ~ slope (Table 6). However, mortality of stems of all species combined was not linked to the environmental variables for which we have data. There was no relationship between mortality and any of the envi- ronmental variables in the case of the two canopy trees D. racemosum and N. aciculata. Patterns of mortality in Eurya japonica, an understory tree (Kohyama 1992), were linked mostly to steeper
Table 5 Diameter at breast height (d.b.h.) of stems that survived Typhoon No. 13, in warm temperate rainforests in the Segire River valley, Yakushima, in various categories of damage caused by the typhoon
Data are presented from two sites: PJB plots (an extensive survey) and TK plots (an intensive survey). Differences between categories of damage were not significant in the TK plots (aNOVa F3,1988 = 0.072, P = 0.98) but were significant in the PJB plots (aNOVa, F4,2156 = 12.89, P = 0.0001; the same letters indicate no significant differences between categories, Scheff6 F-tests, P < 0.05).
slopes (>20 ~ ) and interactions with south-west facing ridge sites.
Severe damage in all species combined was also greatest in higher altitude (>500 m), south-west facing sites (Table 6). Similarly, stems of D. race- mosum were most frequently damaged on higher altitude, south-west facing ridge sites. By contrast, the greatest severe damage to the basal area of E. japonica occurred mostly in areas between south and east on ridges and gullies, and altitudinal effects were not significant. No patterns could be found to relate damage to N. aciculata to the environmental variables for which we have data. Thus, although it is possible to generalize that higher altitude, south- west facing plots had highest levels of damage and mortality, at the individual species level these pat- terns are not so clear. This is due in part to a shortage of data, but some individual species (e.g.E. japon- ica) had different patterns of damage compared with that among all species.
Ordination of common tree species in the PJB plots by Canonical Correspondence Analysis (Fig. 3) showed that their distribution could be explained mostly according to physiography; species with the highest scores on axis 1 were most common on ridge crests and those with the lowest scores in gullies (see Kohyama & Grubb 1994). The effects of increasing altitude are intimately associated with this gradient. Slope was also an important gradient in species distribution, mostly along the second axis. Higher axis 2 scores were typical of species which were most common on steeper sites. Comparing three ordinations based on the extent to which species were killed or damaged (Fig. 3), we inter- preted species with the greatest changes in ordina- tion values to be those most affected by the typhoon. Three species most typically found in lower altitude, less steep sites in gullies (Camellia sasanqua, L. acuminata and P. nagi) showed the least differences between the three ordination scores. However, there was a considerable difference between the ordina- tion score of living basal area and living undamaged basal area for Q. salicina, which was most frequently found in gullies in this survey. There were large differences in ordination scores of Camellia japonica, Illicium anisatum and R. tashiroi, all spedes most typically found in steeper sites such as ridge flanks, but others typically found in these sites such as Cleyera japonica and Myrsine seguinii changed rela- tively little. Although at the level of all species
la ~ . / . S lope Cj
. 2
1 A,wect " �9 ~ L~ ~ A l t i t u d e
Sp =Sp Pnm ~ " I P h " a ' Sb~ yswgr pny
. . . . , . . . . , . . ,
Qs Axis 1 (Eigenvalue = 0.217)
-1 S g ~ e
-2
Fig. 3. Ordination by Canonical Correspondence Ana- lysis of 17 spedes in warm temperate rainforests in an extensive survey (PJB plots) in the Segire River valley, Yakushima, with vectors denoting the influence of four environmental variables. O, Total basal area of stems, living and dead; H, basal area of stems that survived the typhoon; 88, basal area of surviving stems that were not severely damaged by the typhoon. Abbreviations: Cj, Camellia japonica; Cl, Cleyera japonica; Cs; Camellia sasanqua; Dr, Distylium racemosum; Ej, Eurya japonica; Ia, Illicium anisa- turn; La, Litsea acuminata; Ms, Myrsine seguinii; Na, Neolitsea aciculata; Pe, Pasania edulis; Pn, Podocarpus nagi; Qs, Quercus salicina; Rt, Rhododendron tashiroi; Sb, Syzygium buxifolium; Sg, Symplocos glauca; Sp, Symplocos prunifolia; St, Symplocos tanakae.
combined, higher altitude sites were most affected (see earlier), among species most common at higher altitudes (e.g.P. edulis, Symplocos glauca and Syzy- gium buxifolium) there was little change in ordina- tion scores. This analysis suggests that although the degree of damage in some species is to some extent a function of where they grew most frequently, there were also important differences in the effects of the typhoon on individual species which grow in similar sites.
Sprouting
Of the 2161 stems in the PJB plots that survived the typhoon, 375 ( 17.35 %) had epicormic sprouts. Of stems with sprouts, 309 had fresh sprouts and 66 had a 'sprout bank'. There was a considerable range among species in the frequency with which they produced fresh sprouts (Table 4). The frequency of
Tab
le 6
G
ener
al l
inea
r m
odel
s o
f the
rel
atio
nshi
ps b
etw
een
four
env
iron
men
tal
vari
able
s of
the
perc
enta
ge m
orta
lity
and
sev
ere
dam
age
(com
bine
d in
cide
nce
of c
row
n lo
ss,
upro
otin
g an
d se
vere
def
olia
tion
) pe
r pl
ot i
n P
JB p
lots
in
war
m t
empe
rate
rai
nfor
ests
in
the
Seg
ire
Riv
er v
alle
y, Y
akus
him
a
AN
OV
A
Ste
m m
orta
lity
B
asal
are
a m
orta
lity
All
tree
D
isty
lium
E
urya
N
eolit
sea
All
tree
D
isty
lium
E
urya
N
eolit
sea
spec
ies
race
mos
um
japo
nica
ac
icul
ata
spec
ies
race
mos
um
japo
nica
ac
icul
ata
F12,
28 =
1.5
95
F12,
26 =
1.33
1 F1
2,28
= 3,
404
Fll
A9
= 0,
150
F12,
28 =
2.66
2 F1
2,26
= 1.
139
Ft2,
28 =
2.9
51
Fll,1
9 =
0.92
3 P
= 0
.15
P=
0.2
6 P
= 0
.004
P
= 0
.99
P=
0.0
16
P=
0.3
7 P
= 0
.009
P
= 0
.54
Asp
ect
Slop
e A
ltit
ude
Phys
iogr
aphy
A
spec
t x
slop
e A
spec
t x
alti
tude
A
spec
t x p
hysi
ogra
phy
Slop
e x
alti
tude
Sl
ope
• ph
ysio
grap
hy
Alt
itud
e x
phys
iogr
aphy
A
spec
t x s
lope
x a
ltit
ude
Asp
ect x
alt
itud
e x
phys
iogr
aphy
0.59
2.
33*
0.02
2,
47*
2.50
* 2.
10"
1.02
" 2.
87**
1,
48
1.22
1.
18
0.67
2.
69*
2.26
* 2.
20*
0.91
2.
72*
1.37
1.
35
0.80
0.
79
0.26
2.
62*
0.05
2.
82**
0.
52
2.26
* 0.
06
0.89
0.
51
3.09
**
2.25
* 2.
46*
0.09
1.
18
0.50
AN
OV
A
Seve
re d
amag
e to
ste
ms
Seve
re d
amag
e to
bas
al a
rea
All
tree
D
isty
lium
E
urya
N
ed#s
ea
All
tree
D
isty
lium
E
urya
N
eolit
sea
spec
ies
race
mos
um
japo
nica
ac
icul
ata
spec
ies
race
mos
um
japo
nica
ac
icul
ata
F103
0 =
4.14
8 F1
2,2
6 =
2.84
7 F1
2,2
8 =
1.93
6 F
II,1
9 =
1.13
3 Fl
o30
= 2.
300
F12,
2 6
= 1.
319
F12,
2 8
= 2.
162
FIt
,19
= 1.
295
P =
0.03
7 P
= 0.
012
P =
0.07
3 P
= 0.
39
P =
0.03
8 P
= 0.
267
P =
0.04
6 P
= 0.
30
o
o m
Asp
ect
2.17
" 1.
09
1.55
Sl
ope
1.66
O
. 57
0.83
A
ltit
ude
2.91
**
1.88
2.
36*
Phys
iogr
aphy
0.
58
0.34
1,
00
Asp
ect x
slo
pe
2.11
" 0.
73
2.18
" A
spec
t x a
ltit
ude
1.34
1.
83
0.44
A
spec
t x p
hysi
ogra
phy
0.09
O
. 51
0.35
Sl
ope
x al
titu
de
1.04
1.
17
0.40
Sl
ope
x ph
ysio
grap
hy
O. 7
2 1.
07
O. 7
8 A
ltit
ude
x ph
ysio
grap
hy
0.95
2.
07*
1.01
A
spec
t x s
lope
x a
ltit
ude
1.15
A
spec
t x a
ltit
ude
x ph
ysio
grap
hy
2.07
*
2.51
" 0.
67
0,96
2.
16"
1.67
1.
81
2.79
**
1.49
1.
34
1.77
2.
07*
2.18
"
r 0 m g e a
F va
lues
fro
m A
NO
VA
deno
te t
he s
igni
fica
nce
of e
ach
mod
el;
whe
re m
odel
s ar
e si
gnif
ican
t, t
val
ues
and
sign
ific
ance
of
the
effe
cts
of i
ndiv
idua
l en
viro
nmen
tal
vari
able
s an
d k~
inte
ract
ions
am
ong
them
are
sho
wn
(*P
<
0.05
; **
P <
0.
01;
***P
<
0.00
1).
vo
240 P.J. Bellingham et al.
production of fresh sprouts by particular species was strongly correlated with the frequency of severe defoliation by the same species (r17 =0.69, P < 0.001), but was not correlated with frequency of crown loss or uprooting. Of the stems with fresh sprouts, 39% were damaged by the typhoon and the frequency of sprouting of stems damaged in various ways was greater than in comparatively undamaged stems; this pattern was significant for some individ- ual species (Fig. 4). In particular, almost all surviv- ing stems that lost crowns during the typhoon had fresh sprouts. Among all species combined, stems
(a) Stems which lost c rowns
(n = 23) All species combined (n = 2043)
(b) Partially broken s tems
(n : 95) All species combined (n = 2043)
Distylium racemosum (n = 11) (n = 300)
Neolitsea aciculata (n = 13) (n = 125)
(c) Uprooted s tems
(n = 35) All species combined (n = 2126)
(d) SevereIy defoliated s iems
(n = 173) All species combined (n = 1965)
Quercus salicina (n = 15) (n : 2])
Neolitsea aciculara (n = 23) (n = 122)
Symplocosprunifolia (n = 14) (n = 74)
0 20 40 60 80 tOO
Percentage of s tems wi th sprouts
Fig. 4. The percentages of sprouting stems in various damage categories, for all species combined and for individ- ual species. Data are from the extensive survey of forests in the Segire River valley (PJB plots). Differences were compared using adjusted G-tests (n = number of stems in the sample; *P < 0.05; **P < 0.01; ***P < 0.001). (a) i , Crown lost; IZ], crown intact. (b) I , Partially broken; D, crown intact. (c) i , Uprooted stems; D, upright stems. (d) i , Severely defoliated; I-3, not severely defoliated.
> 1 0 c m d.b.h, sprouted more frequently than stems ~ 1 0 cm d.b.h. (G-test, P < 0.05), but there was no difference in the frequency of sprouting of severely damaged stems (i.e. those that lost crowns, were uprooted or severely defoliaced) between the two size classes (G-test, P > 0.05).
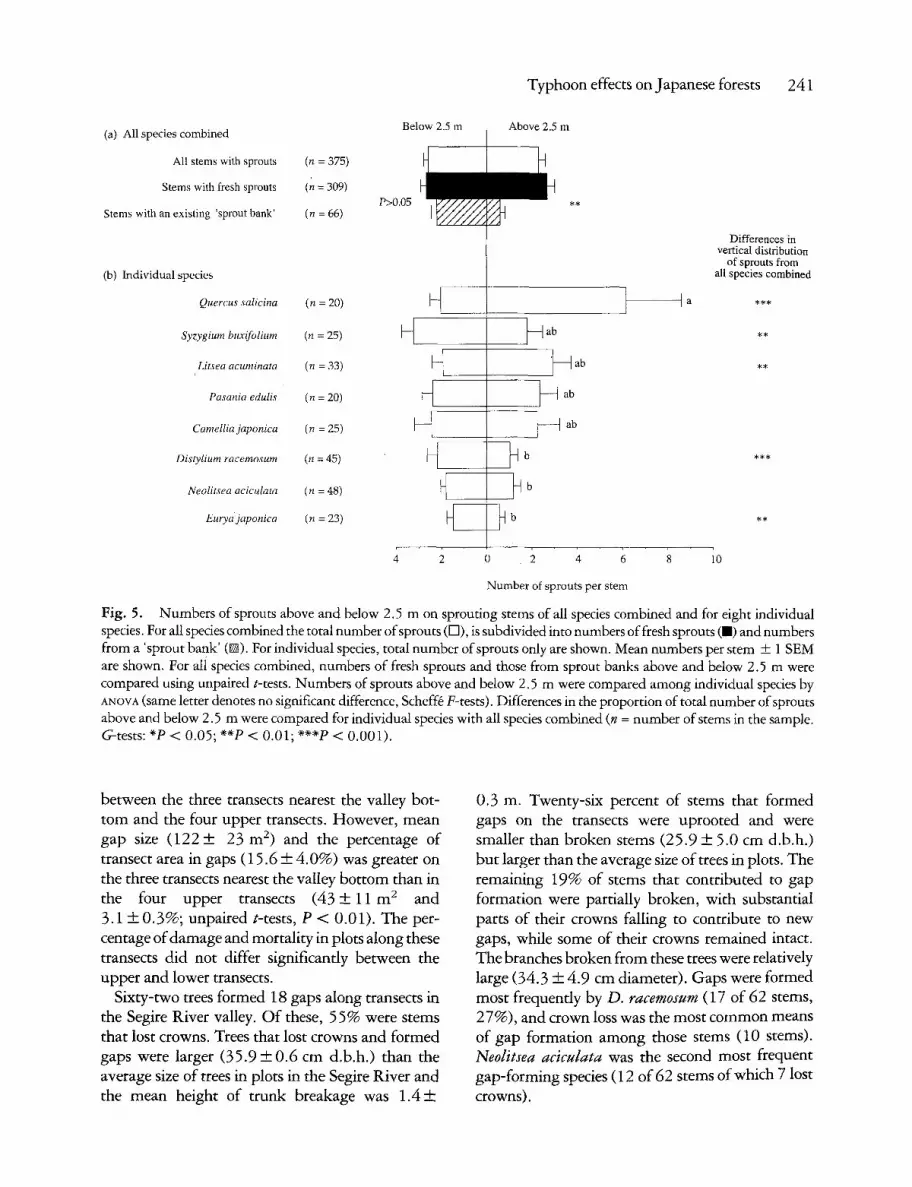
Stems with 'sprout banks' were larger (mean d . b . h . + S E M = 15.4__+2.3 cm) than stems with fresh sprouts (8.5 + 0 . 6 cm) and stems without sprouts (8.2 + 0 . 2 cm); the latter two were not significantly different (aNOVa, F2.2t58 = 17.46, P = 0.0001, Scheff6 F-tests, P < 0.05). Stems with 'sprout banks' had fewer sprouts per stem (mean = 2 . 8 + 0 . 3 ) than stems with fresh sprouts
(5.3 4-0.4). There was no difference in the mean number of sprouts per stem below 2.5 m between stems with 'sprout banks' and those with fresh sprouts (Fig. 5), but above 2.5 m stems with fresh sprouts had more sprouts than did stems with 'sprout banks'. These results suggest that in these forests 'sprout banks' are maintained in relatively small numbers near the bases of larger trunks. Of the 17 common species in Table 4, some stems of all but 4 species (M. seguinii, P. nagi, S. prunifolia and Symplocos tanakae) had 'sprout banks' before Typhoon No. 13.
The mean number of sprouts per stem below 2.5 m was not significantly different for the eight species with 20 or more sprouting stems. By con- trast, the canopy tree Q. salicina produced much greater numbers of sprouts per stem above 2.5 m than the two other canopy trees D. racemosum and N. aciculata (Fig. 5). Distylium racemosum, E. japonica and S. buxifolium had greater proportions of sprouts below 2.5 m than for all species combined, whereas L. acuminata and Q. salicina had greater propor- tions of sprouts above 2.5 m than for all species combined (G-tests, Fig. 5) .
G a p f o r m a t i o n
In the Koyohji River site (0.45 ha), 205 m 2 (4.6%) was in gaps in 1983, but after Typhoon No. 13 in 1993, 424 m 2 (9.4%) was in gap area. In a survey of the valley side in the Segire River catchments along seven transects, 8.6% of the total survey area of 1.77 ha was composed of gaps mostly formed during the typhoon, with a mean frequency ( + SEM) of 2.6 + 0.5 gaps per transect. There was no difference in the number of gaps per transect
Typhoon effects on Japanese forests 241
(a) All species combined
All stems with sprouts (n = 375)
Stems with fresh sprouts ('n = 309)
Stems with an existing 'sprout bank' (n = 66) P>0.05
Below 2.5 m Above 2.5 m
(b) Individual species
Quercus salicina (n = 20)
Syzygium buxifolium (n = 25)
Litsea acuminata (n = 33)
Pasania edulis (n = 20)
Camellia japonica (n = 25) ~ - [
Distylium racemosum (n = 45)
Neolitsea aciculata (n = 48)
Euryajaponica (n = 23) ~ -
4 2
]
~ b
~ ab
' > ab
lab
2 4 6 8 10
Number of sprouts per stem
Differences in vertical distribution
of sprouts from all species combined
l a ***
Fig. 5. Numbers of sprouts above and below 2.5 m on sprouting stems of all species combined and for eight individual species. For all species combined the total number of sprouts (H), is subdivided into numbers of fresh sprouts ( . ) and numbers from a 'sprout bank' (@). For individual species, total number of sprouts only are shown. Mean numbers per stem + 1 SEM are shown. For ali species combined, numbers of fresh sprouts and those from sprout banks above and below 2.5 m were compared using unpaired t-tests. Numbers of sprouts above and below 2.5 m were compared among individual species by aNOVa (same letter denotes no significant difference, Scheff6 F-tests). Differences in the proportion of total number of sprouts above and below 2.5 m were compared for individual species with all species combined (n = number of stems in the sample. G-tests: *P < 0.05; **P < 0.01; ***P < 0.001).
between the three transects nearest the valley bot- t om and the four upper transects. However, mean gap size ( 1 2 2 + 23 m 2) and the percentage of transect area in gaps (15.6 __+ 4.0%) was greater on the three transects nearest the valley bot tom than in the four upper transects ( 4 3 + 1 1 m 2 and 3.1 +__ 0.3%; unpaired t-tests, P < 0.01). The per- centage of damage and mortality in plots along these transects did not differ significantly between the upper and lower transects.
Sixty-two trees formed 18 gaps along transects in the Segire River valley. O f these, 55% were stems that lost crowns. Trees that lost crowns and formed gaps were larger (35 .9-- -0 .6 cm d.b.h.) than the average size of trees in plots in the Segire River and the mean height of t runk breakage was 1 . 4 +
0.3 m. Twenty-six percent of stems that formed gaps on the transects were uprooted and were smaller than broken stems (25.9 + 5.0 cm d.b.h.) but larger than the average size of trees in plots. The remaining 19% of stems that contributed to gap formation were partially broken, with substantial parts of their crowns falling to contribute to new gaps, while some of their crowns remained intact. The branches broken f rom these trees were relatively large (34.3 + 4.9 cm diameter). Gaps were formed most frequently by D. racemosum (17 of 62 stems, 27%), and crown loss was the most common means of gap formation among those stems (10 stems). Ned#sea aciculata was the second most frequent gap-forming species (12 of 62 stems of which 7 lost
c r o w n s ) .
242 P.J. Bellingham et al.
DISCUSSION
Typhoon No. 13 caused low stand-level mortality and damage in our study sites in warm temperate rainforests in Yakushima compared with the effects of many other typhoons in Japanese forests (Table 1). Naka (1982) pointed out that when typhoons affect Japanese coniferous forests, uproot- ing is more frequent than crown loss, which is usually the opposite of forests dominated by angio- sperms such as in our study sites (Table 1). Damage patterns from the Segire River sites are most compa- rable with those from another warm temperate rainforest damaged by a typhoon (Naka 1982), which also contained many species found in our study sites.
Damage and mortality in Yakushima forests caused by Typhoon No. 13 was low compared with that caused by cyclones and hurricanes in other parts of the world (e.g. Unwin et al. 1988; Brokaw & Walker 1991; Elmqvist et al. 1994); its effects may be classified as 'slight' in terms of the classification of hurricane damage devised by Boose et aL (1994). Moreover, the effects of the typhoon on Yakushima forests were variable between catchments only 4.5 km apart (Table 1). The same phenomenon was observed following damage caused by Hurri- cane Hugo to Puerto Rican lower montane rain- forests, where damage recorded in one catchment (Basnet et al. 1992) was much greater than in another less than 10 km away (Walker et al. 1992; Zimmerman etal. 1994). Differences in the ways in which different sites of similar forest composition are affected by cyclones can have important conse- quences for future forest growth (Merrens & Peart 1992).
In Yakushima forests, mortality and non-fatal damage sometimes differed substantially for a spe- cies between two study areas with similar vegetation in the same river valley, less than 1 km apart (Tables 2 and 4). Differences in mortality and damage patterns may also reflect differences in site condi- tions (e.g. soil depth, hence rooting depth) and possible differences in height and wood density for species between the two survey areas, but we lack comparative data. Differences in mortality and damage between the two study sites in the Segire River valley may also be a result of meteorological phenomena during the typhoon, but we lack data to assess how typhoon winds may have varied between
the two sites (cf. Boose et al. 1994). On the valley side, at a stand level, typhoon winds seem to have affected high altitude sites of south-west aspect (Table 6), but these results cannot be generalized to species. The higher incidence of defoliation recorded in these higher altitude, south-west (i.e. seaward) facing sites may have been caused by salt-laden winds. We cannot predict on the basis of this study how future typhoons may affect forests according to topographic position. Some studies have shown that patterns may be predictable at a coarse scale (e.g. Bellingham 1991), but this is not necessarily the case at a fine scale (Zimmerman et al. 1994).
Several studies have shown a range in mortality and damage caused by hurricanes among species (e.g. Wadsworth & Englerth 1959; Walker 1991; Whigham et al. 1991; Elmqvist et al. 1994). The range of non-fatal damage caused by Typhoon No. 13 to tree species in Yakushima forests is lower than in these studies (Tables 2 and 4) and is similar to that found in coastal forests in South Carolina after a hurricane (Gresham et al. 1991). The differences we have found in damage and mortality among species in two sites close by highlight the pitfalls of generalizing from findings based on single events in single sites. However, we do think it is likely that S. [runifatia, which had high mortality in both sites in the Segire River valley, may be more susceptible to the effects of typhoons than other species. Although gymnosperms are often more frequently damaged than angiosperms during hurricanes (e.g. Boucher et al. 1990), the only gymnosperm in the Segire River valley, P. nagi, suffered low mortality and damage during the typhoon in both study sites. When a hurricane affected Jamaican montane forests, an- other species of Podacarpus, P. urbanii, was also relatively little affected (Bellingham et al. 1995). Forests in Yakushima and Jamaica are both affected by cyclonic winds relatively frequently, and natural selection may have resulted in gymnosperm ele- ments more resistant to these winds.
Many studies of the effects of wind in forests have found that larger stems uproot and smaller stems snap (e.g. Putz et al. 1983; Walker et al. 1992). In contrast, in forests in Yakushima, there was no difference between the size of stems that lost crowns and uprooted stems, either living or dead. The same pattern was found in Jamaican montane forests (Bellingham et al. 1995) but Brokaw and Walker (1991) believed this was because of the small range
of diameters of stems in those forests. The lack of differences between the sizes of uprooted and bro- ken stems in Yakushima forests, which have a greater range of stem diameters, may be due in part to the contribution of the common tree D. racema- sum. While large stems of many species in Yakush- ima forests did uproot, large stems of D. racemosum tended to break near their bases rather than uproot. The reason for this may be the high incidence of internal rot (heart rot); many broken stems of this species were hollow.
On Yakushima, Typhoon No. 13 caused defolia- tion in only one of three sites. By contrast, in the Caribbean region, defoliation is the most frequent form of damage caused by hurricanes (Brokaw & Walker 1991). No trees in our study sites were completely defoliated during Typhoon No. 13 in contrast to Jamaican montane forests where Hurri- cane Gilbert completely defoliated 21.2% of stems (Bellinghametal. 1995). However, 8.27% of stems in PJB plots on Yakushima were severely defoliated (i.e. > 50% of leaves lost) during Typhoon No. 13. Our finding that the incidence of severe defoliation of species is negatively correlated with leaf thickness suggests that thinner leaves are expendable in these forests. After a typhoon defoliates trees, there is probably a lower cost of replacement of leaves for thin-leaved species. Although species with thin leaves may be at a competitive disadvantage due to a reduction in leaf area after Typhoon No. 13, this may not be the case after more severe typhoons which may also defoliate thicker-leaved species. When Hurricane Gilbert, which was more powerful than Typhoon No. 13, affected Jamaican montane forests, tree species with the highest spedfic leaf area (SLA) were most frequently completely defoliated (Bellingham et al. 1995), suggesting that these leaves were more brittle and apt to break off than those with a lower SLA.
Most tree species in these forests have population structures which suggest continuous recruitment from seed (reverseJ distributions, Kohyama 1986). While at least three of these species may coexist in these forests on the basis of trade-offs between maximum size and recruitment rates (Kohyama 1992), gap formation not only reduced the time for stable coexistence to be reached for these species but also widened domains of coexistence (Kohyama 1993). After Typhoon No. 13, gap formation appeared to have increased in Yakushima
warm temperate rainforests. In the Koyohji River site, which was less affected than the Segire River sites (Table 1), the gap area after Typhoon No. 13 was more than double that in 1983 before which the forest had not been recently affected by typhoons. Likewise, gap area in a widespread survey of the Segire River valley occupied 8.6% of 1.77 ha, which is nearly double the area of gaps found in a survey in a nearby area long after these forests had last been affected by a typhoon (4.7% of 2.7 ha; Kohyama 1986), although the two survey areas may not be directly comparable. After a typhoon affected warm temperate rainforest in south- western Honshu, Naka (1982) attributed gap for- mation caused by a typhoon as 5.0% of 2.2 ha, but comparisons with the proportion of the forest we surveyed in gap area with this figure may be misleading as we did not feel confident that all recent gaps in our surveys were necessarily directly attributable to Typhoon No. 13. Synchronous cre- ation of sometimes large areas with increased under- story light levels can result from disturbance caused by cyclones in forests, especially where defoliation is widespread (e.g. Turton 1992; Walker et al. 1992). Thus, apparent increases in gap formation and localized defoliated areas resulting from typhoons such as that of 1993 should facilitate coexistence among the 'shade tolerant' species (Kohyama 1993).
After typhoons, opportunities may arise for the regeneration of'pioneer' tree species (sensu Swaine & Whitmore 1988). Yamamoto (1992) found seed- lings of F. ailanthoides, a tree which normally germinates in canopy gaps or at forest margins (e.g. Naka & Yoda 1984), in gaps created during a typhoon in warm temperate rainforest on Tsushima, southern Japan. Five months after Typhoon No. 13, we recorded germination of t7. ailanthoides in Yakushima forests in canopy gaps and also beneath defoliated canopies. Newly germinated seedlings of this species were recorded under defoliated canopies in the PJB plots in densities of 0.67 + 0 . 3 0 m -2. This density is slightly lower than for Chionanthus domingensis but slightly more than for Cecropia schreberiana in similar sites in Puerto Rican lower montane rainforest 5 months after Hurricane Hugo (Guzm~m-Grajales & Walker 1991), but is much lower than for Alchornea latifolia (2000 m -2) in similar sites in Jamaican montane forests 3 months after Hurricane Gilbert (Bellingham et al. 1992).
Although most common tree species in Yaku- shima rainforests are 'shade tolerant', two locally common species, P. edulis and (2. salicina, exhibit distinct 'regeneration gaps' and are rarely recruited from seed, even in canopy gaps (Kohyama et al. 1984; Tanouchi et al. 1994; Yamamoto 1994). During the typhoon, P. edulis suffered high levels of mortality of stems and basal area (Table 2) and both P. edulis and Q. salicina sustained high levels of damage compared with most other species (Table 4). We believe the high incidence of sprout- ing of P. edulis and (2. salicina (often from sprout banks; Fig. 5), and the consequent high incidence of multiple-stemmed individuals, are adaptations that ensure survival of these species in forests frequently disturbed by typhoons. Tanouchi et al. (1994) believed regeneration of (2. salicina from seed or from a seedling bank may follow disturbance to the canopy by severe typhoons. In our study area, we found no existing seedling bank, but direct regener- ation from seed may follow in some sites in future. However, the typhoon struck forests on Yakushima when seeds were probably not fully developed (cf. Tanouchi et al. 1994), so contributions from the current season's seed rain may not be substantial. As conditions which favour their recruitment from seed are rare, we believe the high frequency of sprouting may ensure continued occupancy of sites by these species despite relatively high levels of damage and stem mortality. Bellingham et al. (1994) believed the same was true for some species that seldom regenerate from seed in Jamaican montane forests. Quercus salicina was the species most frequently defoliated during the typhoon (Table 4), but after the typhoon stems of this species produced greater numbers of sprouts higher on stems than other species (Fig. 5). Bellingham et al. (1994) believed that this pattern of sprouting was 'opportunistic' in that these trees could, thus, refill canopy openings. They also found this pattern of sprouting to be usually characteristic of tree species in Jamaican montane forests that mostly regenerate readily from seed, but this is not the case for (2. salicina.
Sprouting after disturbance by cyclonic winds may enable recovery of the canopy mostly by the existing forest trees, a phenomenon described by Yih et al. (1991)as 'direct regeneration'. Particu- larly for stems in which crowns are broken off, sprouting may ensure survival at least in the short term (Bellingham etal. 1994), and in rainforests in
Yakushima, those stems which lost crowns during Typhoon No. 13 and survived did so by sprouting (Fig. 4). After Typhoon No. 13 affected Yakush- ima forests, the overall frequency of sprouting (17.35%) was lower than after hurricanes affected tropical forests on Caribbean islands (61.4% in Jamaican montane rainforests [Bellingham et al. 1994], 64.8% in Puerto Rican lower montane rain forests [Zimmerman et al. 1994]), and while there was a range among species in the frequency with which Yakushima rainforest trees sprouted, the range was less than that reported in these other studies. This may reflect the relatively less severe impact of Typhoon No. 13 on Yakushima forests than the effects of hurricanes in the other studies, or it may reflect biological constraints among the species. For example, P. urbanii sprouted frequently after a hurricane affected Jamaican montane forests (Bellingham et al. 1994), but P. nagi seldom sprouted after the typhoon affected Yakushima forests (Table 4). Other studies in temperate forests affected by wind have shown lower frequencies of sprouting than in tropical forests (e.g. 24.9% in temperate conifer-angiosperm forests in eastern USA [Peterson & Pickett 1991]). Our finding that among both damaged and undamaged stems, larger stems sprouted more frequently than smaller stems is the same as in Jamaican montane rainforests (Bellingham et al. 1994). Several studies have shown that smaller stems severely damaged by wind sprout more readily than larger stems (e.g. Putz et al. 1983; Unwin et al. 1988; Bellingham et al. 1994), but we found no difference in the frequency of sprouting of damaged stems in the two size classes in this study.
In a Jamaican montane forest affected by a hurricane, Bellingham et al. (1995) characterized species in terms of damage (a measure of mortality and frequency of crown loss) and responsiveness (defined by growth rate, recruitment and sprouting frequency). In future we expect to be able to assess responses of Yakushima tree species to the effects of Typhoon No. 13 similarly. However, it is likely that most tree species in Yakushima forests can be considered resistant (i.e. low damage and low re- sponsiveness, especially among the 'shade tolerant' species. In forests that are affected by hurricanes relatively frequently, most tree species seem to be resistant (e.g. Puerto Rican forests, Z immerman et al. 1994; Jamaican forests, Bellingham et al.
1995), thus, it is not surprising that the same pattern should be found in Yakushima forests which are frequently affected by typhoons. In species with higher levels of damage, species such as P. edulis may prove to be resilient (i.e. with high responsive- ness) due to the relative frequency of sprouting, and others such as S. prunifolia due to increased growth rates and recruitment which has been observed in this species in canopy gaps (T. Kohyama, unpubl. data). As a result of generally low levels of mortality and damage caused by the typhoon, we expect that recovery of the forests will occur mostly by 'direct regeneration' of canopy trees. Sprouting of severely damaged stems may ensure at least short-term survival in these forests. However, regeneration from seed and seedling and sapling banks is likely to be important locally, thus, some changes in the relative abundance of species may result from this typhoon.
A C K N O W L E D G E M E N T S
This study received funding from the Japanese Ministry of Education, Science and Culture (Grant No. 93098 and 06304003) and a Japanese Society for the Promotion of Science post-doctoral fellow- ship to PJB. For support at Yakushima we thank F. Iwagawa, K. Harada, N. Akashi and K. Shimano. We also thank the Shimo-Yaku District Forestry Office (Director, Y. Kawano) and the Yakushima Management Office (Head, M. Okuyama) of the Kirishima-Yaku National Park for permission to conduct the study.
REFERENCES
BASNET K., LIKENS G. E., SCATENA F. N. & LuGo A. E. (1992) Hurricane Hugo: damage to a tropical rain forest in Puerto Rico. Journal of Tropical Ecology 8: 47-55.
BELLINGHAM P. J. ( 1991 ) Landforms influence patterns of hurricane damage: evidence from Jamaican montane forests. Biotropica 23: 427-433.
BELLINGHAM P. J., Kayos V., VARXV N. et al. (1992) Hurricanes need not cause high mortality: the effects of Hurricane Gilbert on forests in Jamaica. Journal of Tropical Ecology 8:217-223.
BELEINGHAM P. J., TANNER E. V.J. & HEALEY J. R. (1994). Sprouting of trees in Jamaican montane forests, after a hurricane.Journal of Ecology 82: 747-758.
BELLINGHAM P.J., TANNER E. V.J. & HEALEY J. R. (1995) Damage and responsiveness of Jamaican montane tree species after disturbance by a hurricane. Ecology 76: 2562-2580.
BoosE E. R., FOSTER D. R. & FLUET M. (1994) Hurricane impacts to tropical and temperate landscapes. Ecologi- cal Monographs 64: 369-400.
BOUCHER D. H., VANDERMEER J. H., YIH K. & ZAMORA N. (1990) Contrasting hurricane damage in tropical rain forest and pine forest. Ecology 71: 2022-2024.
BROKAW N. V. L. & WALKER L. R. (1991) Summary of the effects of Caribbean hurricanes on vegetation. Biotro- pica 23: 386-392.
DITTUS W. P. J. (1985) The influence of cyclones on the dry evergreen forests of Sri Lanka. Biotropica 17: 1-14.
ELMQVlST T., RAINEY W. E., PIERSON E. D. & Cox P. A. (1994) Effects of Tropical Cyclones Of a and Val on the structure of a Samoan lowland rain forest. Biotropica 26: 384-391.
FOSTER D. R. (1988a) Species and stand response to catastrophic wind in central New England. Journal of Ecology 76:135-151.
FOSTER D. R. (1988b) Disturbance history, community organisation and vegetation dynamics of the old- growth Pisgah Forest, south-western New Hamp- shire, USA.Journal of Ecology 76: 105-134.
GRESHAM C. A., WILLIAMS T. M. & LIPSCOMB D. J. (1991) Hurricane Hugo wind damage to southeastern U.S. coastal forest tree species. Biotropica 23: 420-426.
GRtSBB P. J., GRUBB E. A. A. & M~YATA I. (1975) Leaf structure and function in evergreen trees and shrubs of Japanese warm-temperate rain forest. I. The structure of the lamina. Botanical Magazine, Tokyo 88: 197- 211.
GUZMXN-GRAJALES S. M. & WALKER L. R. (1991) Differ- ential seedling responses to litter after Hurricane Hugo in the Luquillo Experimental Forest, Puerto Rico. Biotropica 23: 407-413.
INOUE Y. (1959) Forest management (1). In: A Report of Scientific Investigations in Forests Damaged by Wind in 1954, Hokkaido, Japan (eds The Scientific Investiga- tion Group for Wind-Damaged Forests in Hokkaido) pp. 331-436. Japan Forest Technical Association, Tokyo (in Japanese).
KOHYAMA T. (1986) Tree size structure of stands and each species in primary warm-temperate rain forests of southern Japan. Botanical Magazine, Tokyo 99: 267- 279.
KOHYAMA T. (1992) Size-structured multi-species model of rain forest trees. Functional Ecology 6:2196-216.
KOHYAMA T. (1993) Size-structured tree populations in gap-dynamic forest: the forest architecture hypothesis for the stable coexistence of species. Journal of Ecology 81: 131-143.
KOHYAMA T. & GRUBB P. J. (1994) Below- and above- ground allometries of shade-tolerant seedlings in a Japanese warm-temperate rain forest. Functional Eco- logy 8: 229-236.
KOI-IYAMA T., SAKAMOTO K., KOBAYASHI T. & WATANABE R. (1984) Structure of the tree community of a primary lucidophyll forest in the Koyohji basin. In: Conservation Reports of the Yakushima Wilderness Area, Kyushu, Japan (ed. M. Numata) pp. 375- 397. Environment Agency of Japan, Tokyo (in Japanese).
MERRENS E. J. & PEART D. R. (1992) Effects of hurricane damage on individual growth and stand structure in a hardwood forest in New Hampshire, USA.Journal of Ecology 80: 787-795.
M~s~i~Ma T., TANIGUCHI S., TANIGUCHI M. & HISHINUMA Y. (1958) The actual state of wind damage in the Tomakomai Experimental Forest of Hokkaido Uni- versity. II. The natural forest. Research Bulletins of the College Experimental Forests, Hokkaido University 19: 1-39 (in Japanese).
MITSUTA S. & NAGAMASU S. (1984) Vascular plant flora (ferns, fern allies and phanerograms) of the Yaku- shima Wilderness Area. In: Conservation Reports of the Yakushima Wilderness Area, Kyushu, Japan (ed. M. Numata) pp. 103-286. Environment Agency of Japan, Tokyo (in Japanese).
NAKA K. (1982) Community dynamics of evergreen broadleaf forests in southwestern Japan. I. Wind damaged trees and canopy gaps in an evergreen oak forest. Botanical Magazine, Tokyo 95: 385-399.
NAKA K. & YoI)a K. (1984) Community dynamics of evergreen broadleaf forests in southwestern Japan. II. Species composition and density of seeds buried in the soil of a climax evergreen oak forest. Botanical Maga- zine, Tokyo 97: 61-79.
OSAKA REGIONAL FORESTRY OFFICE (1936). A Report on the Typhoon Damage in September 1934. Osaka Regional Forestry Office, Osaka, 179pp. (in Japanese).
PETERSON C. J. & PICKETT S. T. A. (1991) Treefall and resprouting following catastrophic windthrow in an old-growth hemlock-hardwoods forest. Forest Ecology and Management 42: 205-217.
PUTZ F. E., COLEY P. D., Lu K., MONTALVO A. & AIEttO A. (1983) Uprooting and snapping of trees: structural determinants and ecological consequences. Canadian Journal of Forest Research 13:1011-1020.
SHAW W. B. (1983) Tropical cyclones: determinants of pattern and structure in New Zealand's indigenous forests. Pacific Science 37: 405-414.
SHELL D., BURSLEM D. F. R. P., & ALDER D. (1995) The interpretation and misinterpretation of mortality rate measures. Journal of Ecology 8 3 : 3 3 1 - 3 3 3 .
SHIMIZU Y. (1984) The effects of Typhoon No. 17 (6-7 November 1983) on forests in Ogasawara. Ogasa- wara-kenkyu-nenpo 8:21-28 (in Japanese).
SHIMIzu Y. (1994) The ecology and regeneration ofmesic forests in Sekimon, Hahajima Island, Ogasawara Islands: disturbance by Typhoon No. 17 (1983) and recovery processes after the typhoon. Regional Views 7: 3-32 (in Japanese).
SuzuKi E. & TSt:KAHARA J. (1987) Age structure and regeneration of old growth Cryptomeria japonica for- ests on Yakushima Island. Botanical Magazine, Tokyo 100: 223-241.
SWAINE M. D. & WnITMORE T. C. (1988) On the definition of ecological species groups in tropical rain forests. Vegetatio 75: 81-86.
TANOUC~I H., SATO T. & TAKESHITA K. (1994) Compara- tive studies on acorn and seedling dynamics of four Quercus species in an evergreen broad-leaved forest. Journal ofPlant Research 107: 153-159.
TAODA H. (1987) Succession of Castanopsis cuspidata forest accelerated by wind damage. In: Papers on Plant Ecology and Taxonomy in Memory of Dr Satoshi Nakani- shi pp. 379-382. The Kobe Geobotanical Society, Kobe (in Japanese).
TER BRAAK C. J. F. (1986) Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67:1167-1179.
TER BRAAK C. J. F. (1988) CANOCO: an extension of DECORANA tO analyse species-environment relation- ships. Vegetatio 75: 159-160.
ToKyo REO~ONAL FORESTRY OFFICE (1960) Investigations of Wind Damage in Forests Caused by Typhoons No. 7 and No. I5 of I959. Tokyo Regional Forestry Office, Tokyo, 191pp. (in Japanese).
TURTON S. M. (1992) Understorey light environments in a north-east Australian rain forest before and after a tropical cyclone. Journal of Tropical Ecology 8: 241- 252.
UNWrN G. L, AFVLEGATE G. B., STOCKER G. C. & NmHOtSON D. I. (1988) Initial effects of tropical cyclone 'Winifred' on forests in north Queensland. Proceedings of the Ecological Society of Australia 15: 283-296.
WADSWORTH F. H. & ENGLERTH G. H. (1959) Effects of the 1956 hurricane on forests in Puerto Rico. Carib- bean Forester 20:38-51.
WALKER L. R. (1991) Tree damage and recovery from Hurricane Hugo in Luquillo Experimental Forest, Puerto Rico. Biotropica 23: 379-385.
WALKER L. R., VOLTZOW J., ACKERMAN J. D., FERN.~NDEZ D. S. & FETCHER N. (1992) Immediate impact of
Hurricane Hugo on a Puerto Rican rain forest. Ecology 73: 691-694.
WATANABE R., NAKaSHIZUKa T., NONMA S., HaRA M. & YODA S. (1985) Studies on the Fagus crenata forest in Kayanodaira. II. Fallen trees resulting from the winds of Typhoon No. 10 of 1982. Bulletin ofthe Institute for Nature Education, Shiga Heights, Shinshu University 2 2 : 1 5 - 1 8 (in Japanese).
WEAVER P. L. (1986) Hurricane damage and recovery in the montane forests of the Luquillo Mountains of Puerto Rico. Caribbean Journal of Science 22: 53-70.
WEBB S. L. (1989) Contrasting windstorm consequences in two forests, Itasca State Park, Minnesota. Ecology 70: 1167-1180.
WHIGHAM D. F., OLMSTED I., CANOE. C. & HARMON M. E. (1991) The impact of Hurricane Gilbert on trees, litterfall and woody debris in a dry tropical forest in the northeastern Yucatan Peninsula. Biotropica 23: 4 3 4 - 441.
WHITE P. S. (1979) Pattern, process and natural distur- bance in vegetation. Botanical Review 45: 229-299.
WHITMORE T. C. (1974) Change with Time and the Role of Cyclones in Tropical Rain Forest on Kolombangara, Solomon Islands. Commonwealth Forestry Institute Paper 46, Oxford.
WILKINSON L. (1989) SYSTAT: The system for statistics. SYSTAT, Inc. Evanston, Illinois.
YAMAMOTO H., SUZUKI Y., HAYAKAW ̂S. (1994) Agricul- tural damage in Kyushu and Yamaguchi District by Typhoon 9313. Journal of Agricultural Meteorology 49 :285-290 (in Japanese).
YAMAMOTO S. (1992) Gap characteristics and gap regen- eration in primary evergreen broad-leaved forests of western Japan. Botanical Magazine, Tokyo 105: 29- 45.
YAMAMOTO S. (1994) Gap regeneration in primary ever- green broad-leaved forests with or without a major canopy tree, Distylium racemosum, south-western Japan: a comparative analysis. Ecological Research 9: 295-302.
YIH K., BOUCHrR D. H., VANDERMEER J. H. & ZAMORA N. (1991) Recovery of the rain forest of southeastern Nicaragua after destruction by Hurricane Joan. Bio- tropica 23: 106-113.
ZIMMERMAN J. K., EVERHAM E. M. III, WAIDE R. B., LODGE D. J., TAYCOR C. M & BROKaW N. V. L. (1994) Responses of tree species to hurricane winds in sub- tropical wet forest in Puerto Rico: implications for tropical tree life histories. Journal of Ecology 82: 911-922.