The Shape of Rat Kidney Tubular Cells RUTH ELLEN BULGER Department of Anatomy, Harvard Medical School, Boston, Massachusetts ABSTRACT Many cells specialized for ion transport have complex shapes. The cells of the nephron are no exception. Besides the well-documented lateral processes seen in the basal region of many tubular cells, two further specializations are found in rat kidney tubular cells which serve to increase the lateral cell membrane area. These are an interdigitation of processes confined to the apical region, and an inter- digitation of more extensive processes that extend the full height of the cell. Many mammalian renal tubular cells exhibit extreme complexity in shape. Light microscopists of the nineteenth and early twentieth centuries saw and recorded the presence of these extraordinary elabora- tions of cell form. Before 1900, Heiden- hain (1874), Schachowa (1876), Lan- dauer (1895), and Zimmermann (1898) noted the irregular and complicated lateral processes extending from the cells of the proximal convoluted tubule, ascending thick limb of the loop of Henle, and the distal tubules. Zimmermann (1898, '11, '15) identified the complex shape of the cells of the thin limb and visceral glomer- ular epithelium. The application of elec- tron microscopy to the kidney reaffirmed and in some cases extended our under- standing of the configuration of the cells in certain regions of the nephron. Atten- tion has been focused mainly upon elabora- tions of the basal surface of the epithe- lium. In this study, electron micrographs have been obtained which indicate that cell shapes in certain regions of the rat kidney are more complex than has been reported heretofore by electron microscopists. A description is offered of two further com- plexities in the shape of the cells of the renal tubule which serve to increase the area of lateral cell membrane. These are lateral projections in the apical region of the cells and lateral projections that ex- tend the full height of the cells. These findings are in substantial agreement with observations already recorded in the earlier literature of light microscopy, but they re- quire reiteration because they have been overlooked or largely ignored by contempo- rary students of renal fine structure. AM. J. ANAT., 116: 237-256. MATERIAL AND METHODS Kidneys of ten adult rats of the Charles River strain were fixed by three methods: immersion, perfusion, or dripping on the surface of the organ in the living anesthe- tized animal. The fixative used was 1.3% Os04 buffered with s-collidine (Bennett and Luft, '59). In some cases calcium chloride was added to the fixing fluid. Only the superficial regions could be studied in the kidneys processed by drip- ping fixative on the surface. Perfusion was accomplished by injecting cold fixa- tive into the abdominal aorta after clamp- ing the vessel both above and below the level of the renal artery. A vein was cut to allow fluid outflow. The kidneys to be fixed by immersion or after preliminary perfusion were cut into thin pie-shaped pieces with the papilla at the apex and then divided into six regions parallel to the base which were fixed separately to maintain orientation as to the location of the tubules under study. After one hour of fixation, the tissues were rapidly de- hydrated in ethanols and embedded in epoxy resin (Luft, '61). The sections were cut on a Porter-Blum microtome, stained with saturated uranyl acetate and Mil- lonig's lead tartrate ('61), and viewed with RCA3E and 3F microscopes. OBSERVATIONS The complex apical outline of the rat proximal convoluted tubular cell can be seen in a light micrograph of a 1 CI thick section (fig. 1). The terminal bars appear as undulating lines which outline the IPresent address : Department of Pathology, Uniyer- sity of Washlngton Medlcal School, Seattle, Washing- ton 98105. 237
Transcript
The Shape of Rat Kidney Tubular Cells RUTH ELLEN BULGER Department of Anatomy, Harvard Medical School, Boston, Massachusetts
ABSTRACT Many cells specialized for ion transport have complex shapes. The cells of the nephron are no exception. Besides the well-documented lateral processes seen in the basal region of many tubular cells, two further specializations are found in rat kidney tubular cells which serve to increase the lateral cell membrane area. These are an interdigitation of processes confined t o the apical region, and an inter- digitation of more extensive processes that extend the full height of the cell.
Many mammalian renal tubular cells exhibit extreme complexity in shape. Light microscopists of the nineteenth and early twentieth centuries saw and recorded the presence of these extraordinary elabora- tions of cell form. Before 1900, Heiden- hain (1874), Schachowa (1876), Lan- dauer (1895), and Zimmermann (1898) noted the irregular and complicated lateral processes extending from the cells of the proximal convoluted tubule, ascending thick limb of the loop of Henle, and the distal tubules. Zimmermann (1898, '11, '15) identified the complex shape of the cells of the thin limb and visceral glomer- ular epithelium. The application of elec- tron microscopy to the kidney reaffirmed and in some cases extended our under- standing of the configuration of the cells in certain regions of the nephron. Atten- tion has been focused mainly upon elabora- tions of the basal surface of the epithe- lium.
In this study, electron micrographs have been obtained which indicate that cell shapes in certain regions of the rat kidney are more complex than has been reported heretofore by electron microscopists. A description is offered of two further com- plexities in the shape of the cells of the renal tubule which serve to increase the area of lateral cell membrane. These are lateral projections in the apical region of the cells and lateral projections that ex- tend the full height of the cells. These findings are in substantial agreement with observations already recorded in the earlier literature of light microscopy, but they re- quire reiteration because they have been overlooked or largely ignored by contempo- rary students of renal fine structure.
AM. J. ANAT., 116: 237-256.
MATERIAL AND METHODS
Kidneys of ten adult rats of the Charles River strain were fixed by three methods: immersion, perfusion, or dripping on the surface of the organ in the living anesthe- tized animal. The fixative used was 1.3% Os04 buffered with s-collidine (Bennett and Luft, '59). In some cases calcium chloride was added to the fixing fluid. Only the superficial regions could be studied in the kidneys processed by drip- ping fixative on the surface. Perfusion was accomplished by injecting cold fixa- tive into the abdominal aorta after clamp- ing the vessel both above and below the level of the renal artery. A vein was cut to allow fluid outflow. The kidneys to be fixed by immersion or after preliminary perfusion were cut into thin pie-shaped pieces with the papilla at the apex and then divided into six regions parallel to the base which were fixed separately to maintain orientation as to the location of the tubules under study. After one hour of fixation, the tissues were rapidly de- hydrated in ethanols and embedded in epoxy resin (Luft, '61). The sections were cut on a Porter-Blum microtome, stained with saturated uranyl acetate and Mil- lonig's lead tartrate ('61), and viewed with RCA3E and 3F microscopes.
OBSERVATIONS
The complex apical outline of the rat proximal convoluted tubular cell can be seen in a light micrograph of a 1 CI thick section (fig. 1). The terminal bars appear as undulating lines which outline the
IPresent address : Department of Pathology, Uniyer- sity of Washlngton Medlcal School, Seattle, Washing- ton 98105.
237
238 RUTH ELLEN BULGER
apical interdigitation of the cells in that part of the nephron. In addition to small secondary irregularities or extensions, larger lateral processes can be seen. These correspond to the specializations in cell shape that are the subject of this study of rat kidney tubular cells.
The first consists of an extensive inter- locking of fine processes that extend ra- dially from the cell apex. In electron micrographs, small cytoplasmic compart- ments are frequently observed at the lu- minal border of the epithelium (figs. 2, 3 ) . These enclaves of cytoplasm are bounded by a continuous plasma membrane and joined to neighboring cells by typical junc- tiond complexes. They are interpreted as cross sections of the lateral processes that extend radially from the apical portion of the epithelial cells and evidently corre- spond to the fine processes whose inter- digitations are seen as the fine irregulari- ties of the boundary contour in views from the luminal surface with the light micro- scope. In sections of proximal convoluted tubules that pass tangential to the lumen, the continuity of these processes with the cell body can be seen and the complexity of this apical interdigitation can be more fully appreciated (fig. 4). These apical interdigitating processes are most numer- ous in the proximal convoluted tubule and in the upper portion of the thin limb of the loop of Henle. They are less abundant in the ascending thick limb and the distal tubule. Such apical extensions have been seen in the proximal convoluted tubule of dog, cat, gerbil, and human (Bulger, un- published observation). The emphasis in previous electron microscopic studies has been upon amplification of surface at the cell base. The apical interdigitation seen in these species serves to increase the area of lateral cell membrane in the apical region of the cells.
The second category of surface special- ization to which we wish to draw a.ttention is lateral compartments of varying width, that are not confined to the apical region, but extend the full height of the epithelium from lumen to base. They seem to be radial processes similar in all respects to those already described for the squamous cells of the loop of Henle (Rhodin, '58a,b). They are connected to neighboring cells by
junctional complexes and differ from the apical processes described above and from the basal lateral processes of Clark ('57) and Rhodin ('58a,b) only in that they extend from lumen to base. They are most numerous in the first part of the ascending thick limb and in the thin limb (Rhodin, '58a,b) but occur also in the proximal convoluted tubule and in limited numbers in the cortical part of the ascending limb, and distal convoluted tubule. An electron micrograph is presented in figure 5 of the proximal part of the ascending thick limb in the inner stripe of the rat medulla. Many compartments are seen bounded by lateral cell membranes that have junctional complexes near the luminal border. In this region of the nephron, the compart- ments are often only wide enough to hold one mitochondrion. A similar region is shown in figure 6. Processes of the cell at the bottom of the figure can be seen to alternate with processes of other cells that extend from base to lumen hence indica- ting a similar pattern of lateral interdigita- tion.
A low power electron micrograph of the proximal tubule (fig. 7) demonstrates both of these categories of surface elaboration. The long arrows on the figure mark junc- tional complexes where the cell membrane is seen to run directly to the basal surface - hence would normally be interpreted as lateral boundaries of cells. The large num- ber of these membrane pairs fa r exceeds the number of cells comprising the cross section of the tubule and hence gives some measure of the abundance of slender lat- eral processes that extend from base to lumen. The shorter arrows mark junc- tional complexes where the cell mem- branes run from the lumen into the cell for a short distance and return to the surface - demarcating small lateral proc- esses that are confined to the apical region of the cell. A schematic three-dimensional representation of the rat proximal convo- luted tubular cell is presented in figure 8 showing apical interdigitation, lateral in- terdigitation, and basal-lateral interdigita- tion.
The two specializations described here along with the well-documented lateral processes of the basal region of the cell all
KIDNEY CELL SHAPE 239
increase the area of lateral cell membrane contact.
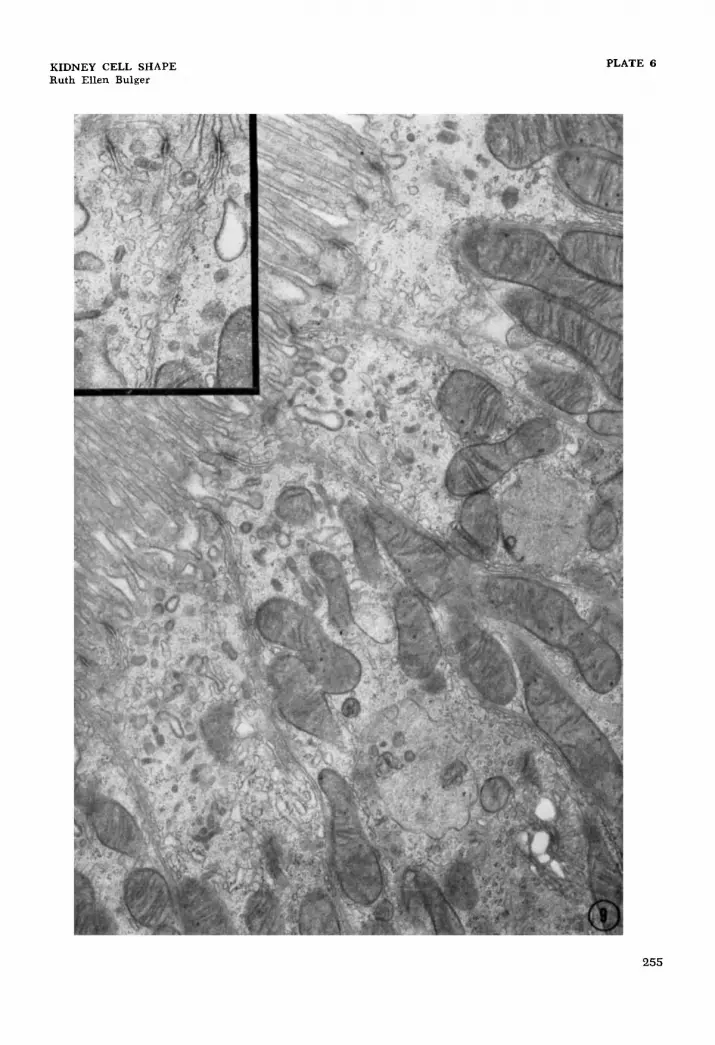
Elements of the smooth-surfaced endo- plasmic reticulum are very commonly in close apposition to the lateral cell mem- branes, and because this relationship may have some functional significance in rela- tion to the amplification of lateral cell sur- face, it will be described here. This special localization of the smooth-surfaced mem- branes was first noted in an abstract by Ericsson et al. (’63). It was visualized by these authors as a continuous tubular sys- tem extending from the apical region to the middle and basal parts of the rat proxi- mal convoluted tubular cell. In our prep- arations, the smooth-surfaced profiles appear as an anastomosing system of ir- regularly shaped elements (fig. 9). This system can be seen along all the lateral cell membranes : those bounding apical compartments, those running from the lumen to base, and those surrounding the lateral processes seen in the basal cyto- plasm. The system is most highly devel- oped along the membranes in the apical part of the cell although it can extend throughout the height of the cell. The Golgi apparatus often lies close to the smooth - surfaced membranes described above. This may reflect some functional association between the two but, on the other hand, it should be kept in mind that the large number of lateral interdigitations in the proximal convoluted tubular cells increase the probability of seeing the smooth-surfaced membranes in close spa- tial relation to the Golgi apparatus.
DISCUSSION
The remarkable elaboration of cell out- lines in the renal epithelium which is now commonly included among the contribu- tions of electron microscopy was actually clearly established by the work of several discerning light microscopists from 1874 to 1942. In 1874, Heidenhain found that the cytoplasm of the mammalian proximal convoluted tubule, ascending thick limb of the loop of Henle, and the distal tubules contained a large number of little rods which he called “Stabchen.” Isolated cells from the dog proximal convoluted tubules were depicted by him as having large arms of cytoplasm that extend laterally and give
off small secondary processes. Schachowa (1876) discussed the complicated lateral borders of the basal portion of dog kidney tubular cells. Although these interdigita- ting grooves and furrows varied in shape and number, they were seen in the cells of the proximal convoluted tubule, thick ascending limb, and distal tubule. Similar flutings of the lateral cell surface were seen in several mammalian species (including human) by Landauer (1895). He estab- lished that the radiating processes existed in the uppor portion of the cell as well as at the base. Using cat and dog kidneys, Zimmermann (1898, ’11, ’15) confirmed the earlier studies. He stressed the transi- tion between the complex apical outline of the pars convoluta and the simple apical outline of the pars recta. The extremely complex form of the cells of the thin limb and the visceral glomerular epithelium were identified and beautifully illustrated by Zimmermann. A comprehensive review of these and additional investigations was written by von Mollendorff in 1930.
Graf€lin and Foote (’38, ’39) and Foote and Graffiin (’42) studied the two seg- ments of the proximal tubule in the cat and dog nephron by silver impregnation of the highly complicated cell boundaries. The cells of the first segment of the proxi- mal tubule of the cat were found to be extraordinarily interdigitated at the sur- face, less complex in the intermediate re- gion, and again extremely irregular near the basement membrane. Those of the second segment of the proximal tubule were essentially rectilinear at the luminal surface but usually interdigitated in vari- able degree near the basement membrane. In the first segment of the proximal tubule of the dog nephron, the cell shapes were reported to be far more irregular than those of the cat and no significant differ- ences in the complexity of their outline could be detected at the luminal, inter- mediate, and basal levels of the cells. Chrome-silver preparations of the second segment of the dog proximal tubule are similar to those described in the cat but the simple rectilinear cell outline seems to be present from lumen to basement mem- brane.
Early electron microscopic studies of the kidney in some cases extended but in
240 RUTH ELLEN BULGER
others actually confused the understand- ing of cell shapes in the nephron. Pease and Baker (’50) regarded the fine processes on cells of the visceral layer of Bowman’s capsule as ridges of the basement mem- brane instead of cellular processes. In sec- tions subtangential to the tubular lumen, they showed the interlocking of the basal regions of the proximal tubular cells. In transverse sections, however, cutting the cells from basement membrane to lumen, the numerous cell membranes at the base of the proximal and distal tubular epi- thelia were not clearly resolved and were interpreted as condensations of cytoplasm forming tubular sheaths around the mito- chondria. In 1951 Dalton recognized that the cells of the visceral layer of Bowman’s capsule have fine cytoplasmic processes. He also pointed out the tortuous lateral cell membranes in a proximal tubular cell but then failed to appreciate the implications of the irregularity of outline for interpreta- tion of sections in the other plane, and simply described basally located intracell- ular “filament-like structures” in both proximal and distal tubular cells.
Sjostrand and Rhodin (’53) noted the basal cytoplasm of the proximal tubules of mice to be divided into open compartments by well-defined intracellular double mem- branes. The compartments were thought to be closed basally, opening into the peri- nuclear or intermediate zone of cytoplasm. In 1954 Rhodin proposed that the intra- cellular membranes seen in thin sections of the basal region of the cytoplasm repre- sented a complicated system of infoldings of the basal cell membrane. According to this interpretation, the folds of cell mem- brane delimit narrow parallel compart- ments of the overlying cell that are open to the cell body above and extend down- ward to the basement membrane. Pease (’55a,b, ’56), in his later papers, agreed that the basal region of the cells of the proximal convoluted tubule was subdivided by ‘infoldings or intussusceptions” of the plasma membrane. He drew attention to the fact that this specialization was even more elaborate in the distal tubule. Ruska et al. (’57) noted that the “infolded” cell membranes rarely, if ever, return directly to the cell base. The membranes generally return to the basement membrane at a
distance from the point of infolding and enclose portions of cytoplasm. They de- scribe a labyrinthine system of intercom- municating potential spaces between the paired membranes at the base of the epi- thelium.
As a result of studies of cellular dif- ferentiation in the kidneys of newborn mice, Clark (’57) seems to have acquired a clear understanding of the shapes of the cells. He found that at birth the basal cell membrane was relatively simple in contour while the lateral cell membrane was often tortuous and interlocking. The formation of the basal striations which be- gins after birth was interpreted to be the result of progressive flutings of the col- umnar cells. In two extensive papers pub- lished in 1958, Rhodin concluded that the basal parts of the proximal and distal con- voluted tubular cells are provided with a multitude of radially disposed thin plate- like cytoplasmic processes that interlock with those of neighboring cells like the meshing of cogwheels. The sides of the upper portion of the proximal tubular cells were said to be smooth contoured without any radiating lamellar processes. In later reviews, Rhodin (’62, ’63) continued to diagram the cells of the proximal convo- luted tubule with rectilinear cell borders in the apical part of the cell.
Many electron microscopic studies have focused attention on specializations of the basal cell membrane in epithelia special- ized for ion transport. Pease (’56) noted the vast increase and postulated that such a modification might increase the area of active surface for ion transport. Similar modifications were discovered that in- creased the membrane area on the lateral surface of cells in many of these transport- ing tissues (Millen and Rogers, ’56; Max- well and Pease, ’56; Pease, ’56; Pappas and Smelser, ’58, ’60, ’61; and Munger, ’61). Some tissues such as gall bladder (Ya- mada, ’55; and Hayward, ’62a,b) have membrane specializations almost entirely on the lateral surface.
From recent work, including that of Clark (’57) and Rhodin (’58a, b), it has become increasingly apparent that many of the structures first identified as basal infoldings are in fact interdigitating lat- eral processes occupying corresponding re-
KIDNEY CELL SHAPE 24 1
cesses in adjacent cells. Parks ('61) re- discovered that the basal region of the cells of the striated duct of mouse parotid gland has leaf-like processes which fit into grooves on adjacent cells. While studying the ciliary epithelium of the adult albino rabbit, Tormey ('63) found that the "in- folded plasma membranes" described by previous workers were cell membranes covering projections or interdigitations from adjacent cells. By studying serial sections, he showed that the apical (em- bryologically basal) and lateral interdigita- tions appear to be two aspects of the same structure. Hence, many surface special- izations which were formerly thought to be mechanisms for increasing the basal cell membrane area, in fact, increase the area of lateral cell surface.
Various amounts of lateral interleafing exist between cells of non-mammalian or- gans involved in the movement of ions. The dogfish rectal (salt-secreting) gland consists of cells with apical interdigita- tions and lateral projections (Bulger, '61, '63). Christensen ('63) has shown that the distal tubular cells of the salamander kidney (Batrachoseps attenuatus) have thin lamellae which interdigitate with ad- jacent cells. The cells of the nasal salt- secreting gland of the lizard (Dipsosaurus dorsalis) have lateral infolded processes (Philpott and Templeton, '64). Extreme complexities in cell shape have been re- ported in the nasal gland of marine birds by Doyle ('60a,b) and in the calciferous gland of the earthworm (Lumbricus ter- restris) by Ellis ('63).
The cells throughout the rat nephron tend to have a branching, star-shaped form. In some regions of the tubules, the irregular outlines frequently extend the entire height of the cell as in the proximal convoluted tubule, the squamous thin limb cells, and the first part of the ascending thick limb. In other regions the branching form involves mainly the basal region of the cells as in the latter part of the thick ascending limb and the distal convoluted tubule. The visceral epithelium of the glomerulus also has many radiating and branching processes giving rise to the familiar pedicels.
It is difficult and probably premature to attempt to assign any specific function
to the complex shape of the rat kidney cells. However, certain observations seem appropriate. The form of the kidney cells leads to an increase in the lateral cell sur- face area, an increase in the intercellular space, and an increase in area of cell to cell contact. Since much of the kidney tubule is involved in ion transport proc- esses, it is probably safe to assume that the complex cell shape is related to these processes. This is further substantiated by the fact that cells with complex shapes are found in other organs specialized for ion transport in mammalian and non- mammalian species. However, some doubt is cast on this interpretation by the fact that the branching cell shape is found in two regions of the nephron not thought at present to be involved with active ion transport : the visceral epithelium of Bow- man's capsule (podocyte) and the descend- ing thin limb of the loop of Henle. The interleafing processes of the podocyte may offer support while giving rise to a large number of filtration slits. Although it has been postulated that the cells of the as- cending portion of the loop of Henle (in- cluding the thin limb cells) contain an active pump which moves sodium into the interstitium of the papilla thus setting up the countercurrent multiplier system, the descending limb is thought to play only a passive role. No distinct morphological differences have been demonstrated be- tween the cells of the ascending and de- scending limbs in spite of the seemingly different physiological roles.
The smooth-surfaced anastomosing sys- tem of membrane limited elements found along the lateral cell membrane may be functionally related to the lateral cell mem- brane. Ericsson et al. ('63) postulated that the part of the endoplasmic reticu- lum associated with the lateral cell mem- branes may represent a transport system connecting the apical cytoplasm with the intercellular space and the Golgi-cytosome region. Although as yet there is no phys- iological data to support this hypothesis, the close morphological association of these elements is at least circumstantial evidence for some functional interaction between the cytoplasmic and surface mem- branes.
242 RUTH ELLEN BULGER
ACKNOWLEDGMENTS
The author wishes to express her thanks to Dr. Don Fawcett for the opportunity to do this work and for his continued en- couragement throughout the course of the study.
This investigation was supported by USPHS grant GM-10182 and NSF grant G-129 16.
LITERATURE CITED
Bennett, H. S., and J. H. Luft 1959 s-Collidine as a basis for buffering fixatives. J. Biophysic. Biochem. Cytol., 6: 113-114.
Bulger, R. E. Unpublished observations. 1961 Histology and ultrastructure of
the rectal gland of Squalus sucklii (acanthhs) . First Annual Meeting of the American Society for Cell Biology, 24 (abstract). - 1963 Fine structure of the rectal (salt-
secreting) gland of the spiny dogfish, Squalvs acanthias. Anat. Rec., 147: 95-127.
Christensen, A. K. 1963 Fine structure of an exceptional kidney in the salamander, Batro- choseps. J. Cell Biol., 19: 13A (abstract).
Clark, S. L., Jr. 1957 Cellular differentiation in the kidneys of newborn mice studied with the electron microscope. J. Biophysic. Biochem.
Dalton, A. J. 1951 Structural details of some of the epithelial cell types in the kidney of the mouse as revealed by the electron microscope. J. Natl. Cancer Inst., 11: 1163-1186.
Doyle, W. L. 1960a The principal cells of the salt gland of marine birds. Exper. Cell Res.,
Fine structure of salt regulating epithelia. Anat. Rec., 136: 184 (abstract).
Ellis, R. A. 1963 The fine structure of the secretory epithelium in the calciferous glands of the earthworm. Anat. Rec., 145: 226 (abstract).
Ericsson, J., F. Mostofi and J. Weibel 1963 Ultrastructure of proximal convoluted tubules : endoplasmic reticulum, Golgi apparatus and single membrane limited bodies. Anat. Rec., 145: 227 (abstract).
Foote, J. J., and A. L. Grafflin 1942 Cell con- tours in the two segments of the proximal tu- bule in the cat and dog nephron. Am. J. Anat., 70: 1-20.
Grafflin, A. L., and J. J. Foote 1938 The Bodian technique as applied to the complicated cell boundaries of the renal epithelium in mam- mals. Anat. Rec., 72: 115-118.
Epithelial cell shapes in the first segment of the proximal tubule of the cat nephron as demonstrated by the chrome-silver method. Am. J. Anat., 65: 179-198.
Hayward, A. F. 1962a Electron microscopic ob- servations on absorption in the epithelium of the guinea pig gall bladder. Zeitsch. Zellforsch.,
Cytol., 3: 349-362.
21: 386-393. 1960b
1939
56: 197-202.
1962b Aspects of the fine structure of the gall bladder epithelium of the mouse. J. Anat., London, 96: 227-236.
Heidenhain, R. 1874 Mikroskopische Beitrage zur Anatomie und Physiologie der Nieren. Arch. f. Mikr. Anat., 10: 1-50.
Landauer, Armin 1895 Ueber die Structur des Nierenepithels. Anat. Anz., 10: 645-653.
Luft, J. H. 1961 Improvements in epoxy resin embedding methods. J. Biophysic. Biochem.
Maxwell, D. S., and D. C. Pease 1956 The electron microscopy of the choroid plexus. J. Biophysic. Biochem. Cytol., 2: 467-474.
Millen, J. W., and G. E. Rogers 1956 A n elec- tron microscopic study of the choroid plexus in the rabbit. J. Biophysic. Biochem. Cytol., 2: 407-416.
Millonig, G. 1961 A modified procedure for lead staining of thin sections. J. Biophysic. Biochem. Cytol., 11: 736-739.
Mollendorff, W. von 1930 Handbuch der Mikro- skopischen Anatomie des Menschen. Siebenter Band. Harn und Geschlechtsapparat. Berlin.
Munger, B. L. 1961 The ultrastructure and histophysiology of human eccrine sweat glands. J. Biophysic. Biochem. Cytol., 11: 385402.
Pappas, G. D., and G. K. Smelser 1958 Studies on the ciliary epithelium and the zonule. Am. J. Ophthalmol., 46: 299-318.
1960 The fine structure of the ciliary epithelium in relation to aqueous secretion. Anat. Rec., 136: 254-255 (abstract).
1961 The fine structure of the ciliary epithelium in relation to aqueous humor secre- tion. In: The Structure of the Eye. Ed. G. K. Smelser, Academic Press, New York, 453-467.
Parks, H. F. 1961 On the fine structure of the parotid gland of mouse and rat. Am. J. Anat., 108: 303-329.
Pease, D. C. 1955a Fine structure of the kid- ney seen by electron microscopy. J. Histochem. Cytochem., 3: 295-308.
1955b Electron microscopy of the tubular cells of the kidney cortex. Anat. Rec., 121: 723-743.
1956 Infolded basal plasma mem- branes found in epithelia noted for their water transport. J. Biophysic. Biochem. Cytol., 2
Pease, D. C., and R. F. Baker 1950 Electron microscopy of the kidney. Am. J. Anat., 87: 349-390.
Philpott, C. W., and J. R. Templeton 1964 A comparative study of the histology and fine structure of the nasal salt secreting gland of the lizard, Dipsosaums. Anat. Rec., 148: 394 (abstract).
Rhodin, J. A. G. 1954 Correlation of ultra- structural organization and function in normal and experimentally changed proximal convo- luted tubule cells of the mouse kidney. Thesis, Stockholm, Sweden.
1958a Electron microscopy of the kid- ney. Am. J. Med., 24: 661-675.
1958b Anatomy of kidney tubules. Intern. Rev. Cytol., 7: 485-534.
Cytol., 9: 409-414.
( S U ~ P ~ ) : 203-208.
KIDNEY CELL SHAPE 243
1962 Electron microscopy of the kid- ney. In: Renal Disease. Ed. by D. A. K. Black, Blackwell Scientific Publications, F. A. Davis Company, Philadelphia, Pennsylvania.
1963 Structure of the kidney. In: Dis- eases of the Kidney. Ed. by M. B. Strauss and L. G. Welt, Little, Brown and Company, Boston, Massachusetts.
1957 The base of the proximal convoluted tubule cells of the rat kidney. J. Biophysic. Biochem.
Schachowa, S. 1876 Untersuchunger uber die Nieren. Dissertation, Bern. As quoted by von MoUendorfF.
Sjostrand, F. S., and J. A. G. Rhodin 1953 The ultrastructure of the proximal convoluted tu- bules of the mouse kidney as revealed by high
Ruska, H., D. H. Moore and J. Weinstock
Cytol., 3: 249-254.
resolution electron microscopy. Exp. Cell Res., 4: 426456.
Tormey, J. 1963 Fine structure of the ciliary epithelium of the rabbit, with particular refer- ence to “infolded membranes,” “vesicles,” and the effects of diamox. J. Cell Biol., 17: 641- 659.
Yamada, E. 1955 The fine structure of the gall bladder epithelium of the mouse. J. Biophysic. Biochem. Cytol., 1: 445458.
Zimmermann, K. W. 1898 Beitrage zur Kennt- niss einiger Drusen und Epithelien. Arch. mikr. Anat., 52: 552-706.
191 1 Zur Morphologie der Epithelzellen der Saugetierniere. Arch. mikr. Anat., 78:
1915 Uber das Epithel des glomeru- laren Endkammerblattes der Saugerniere. Anat. Anz., 48: 335-341.
199-231.
PLATE 1 EXPLANATION OF FIGURES
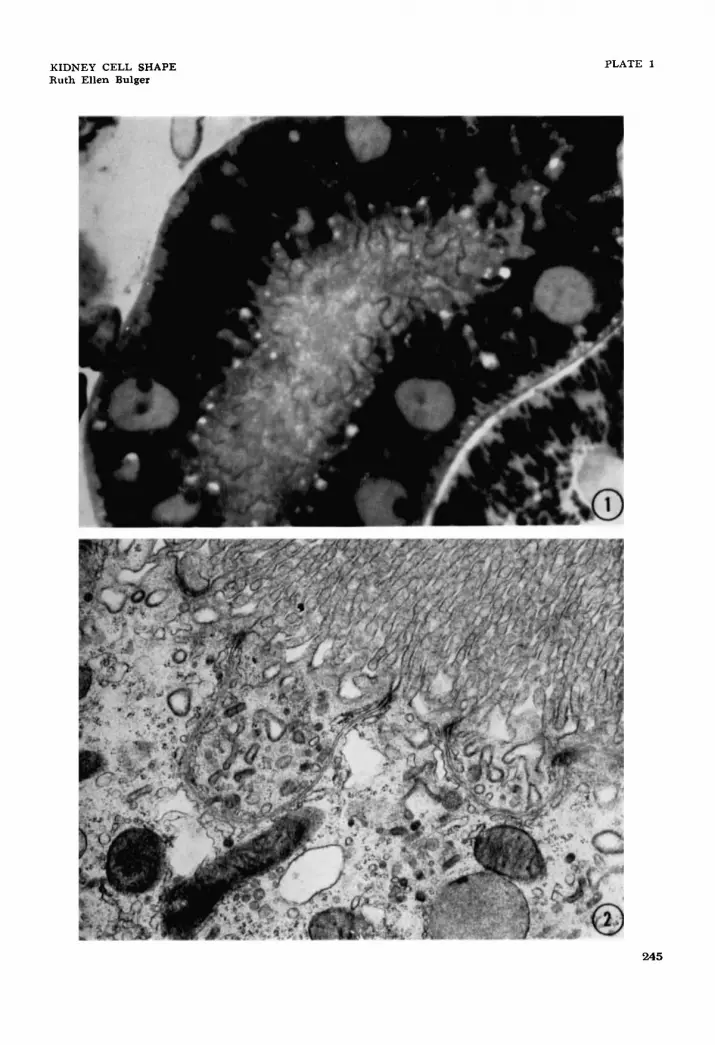
1 Light micrograph of a 1 p thick Epon section, stained with toluidine blue, of a rat proximal convoluted tubule cut obliquely. The terminal bars can be seen as undulating lines that outline the complex inter- digitations of the cells in this part of the nephron. There are large processes as well as smaller irregularities. X 2,400.
Electron micrograph of the apical region of a proximal convoluted tubular cell of the rat. Small cytoplasmic compartments are seen, bounded by continuous plasma membrane and joined to neighbor- ing cells by junctional complexes, which are interpreted as lateral extensions of the apical region of neighboring cells. x 21,600.
2
244
KIDNEY CELL SHAPE Ruth El len Bulger
PLATE 1
245
PLATE 2
EXPLANATION OF FIGURES
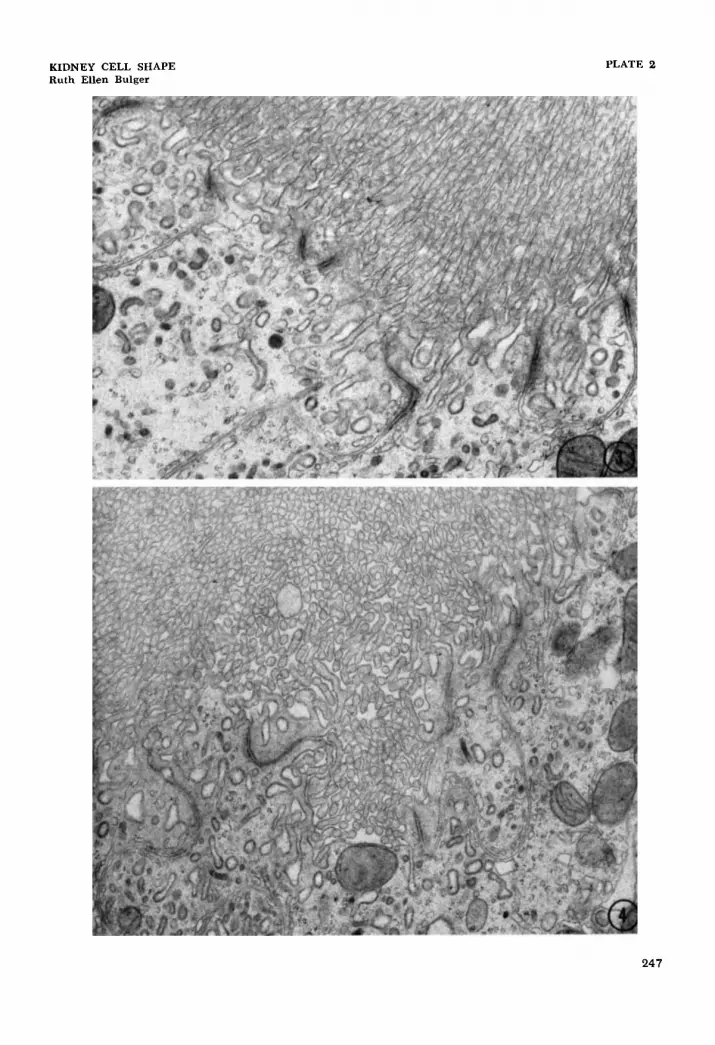
3 Electron micrograph showing three compartments in the apical cyto- plasm. The lateral cell membrane dividing the two cells has a tortuous course. x 19,400.
4 The apical interdigitating processes can be followed in this some- what tangential section. The cell a t the lower left can be seen to abut on the lumen in four places. X 16,800.
246
KIDNEY CELL SHAPE Ruth Ellen Bulger
PLATE 2
247
PLATE 3
EXPLANATION OF FIGURES
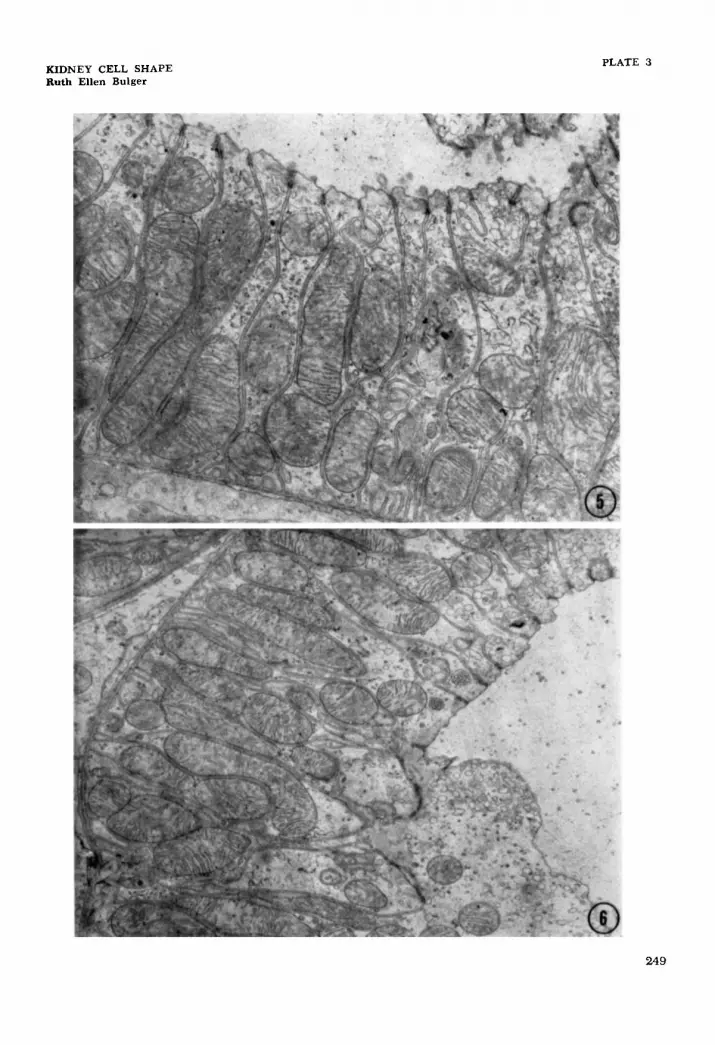
5 Electron micrograph of early part of the ascending thick limb located in the inner stripe of the rat medulla. Many compartments are seen separated by lateral cell membranes which are interpreted as profiles of lateral processes that extend for the height of the cell. Examples of lateral compartments in the apical region of the cell can also be seen. x 12,500. Electron micrograph of early part of the ascending thick limb located in the inner stripe of the rat medulla. The dense areas identify terminal bar regions. The cytoplasm of the cell on the bottom can be seen to interdigitate with the lateral processes of what are presumed to be other cells. x 8,200.
6
248
KIDNEY CELL SHAPE Ruth Ellen Bulger
PLATE 3
249
PLATE 4
EXPLANATION OF FIGURE
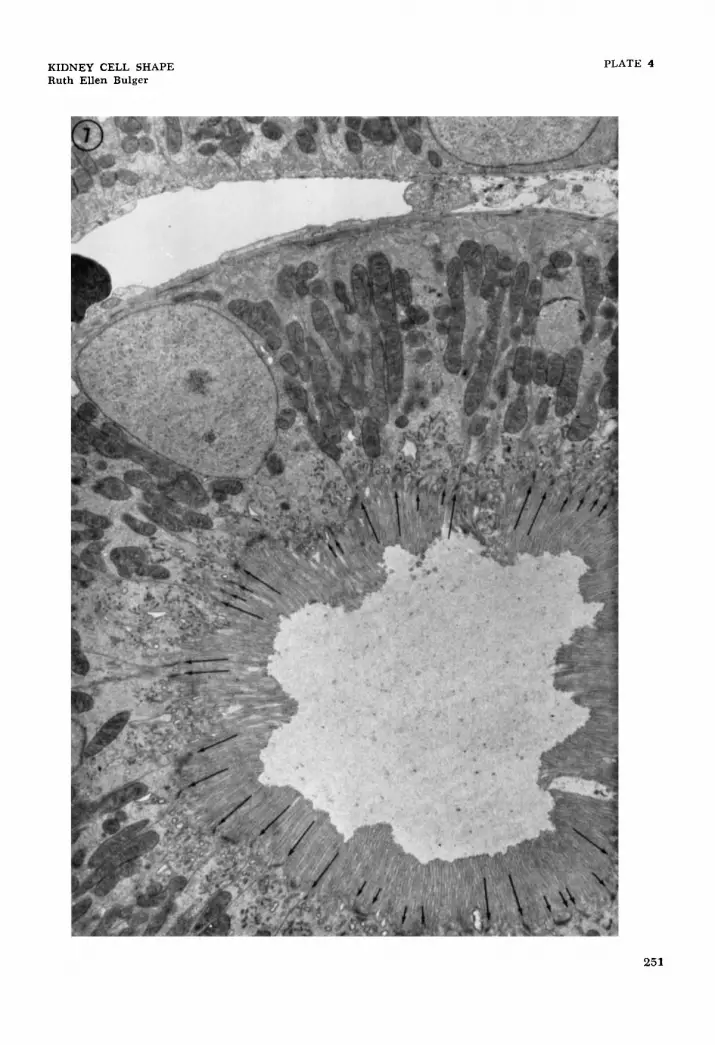
7 Electron micrograph of a rat proximal convoluted tubule. The long arrows mark junctional complexes where the cell membrane seems to run directly to the basal surface - hence should be lateral bound- aries of cells. The large number of these lateral boundaries indicate lateral processes which extend from base to lumen. The short arrows mark junctional complexes where the cell membranes run into the cell for a short distance and return to the surface - demarcating small lateral processes in the apical region of the cell. x 5,600.
250
KIDNEY CELL SHAPE Ruth Ellen Bulger
PLATE 4
251
PLA
TE
5
EX
PL
AN
AT
ION
O
F
FIG
UR
E
8
A d
iagr
am o
f ra
t pr
oxim
al c
onvo
lute
d tu
bula
r ce
lls
show
ing
thre
e ty
pes
of
late
ral
inte
rdig
itat
ing
proc
esse
s :
thos
e in
the
api
cal
regi
on,
thos
e in
the
bas
al
regi
on,
and
tho
se
exte
ndin
g fo
r th
e he
ight
of
th
e ce
ll.
KID
NE
Y C
EL
L S
HA
PE
R
uth
Ell
en B
ulge
r PL
AT
E 5
PLATE 6
EXPLANATION OF FIGURE
9 Electron micrograph of rat proximal convoluted tubule. Elements of the smooth-surfaced endoplasmic reticulum are commonly seen in close apposition to the lateral cell membranes. X 15,000. The inset shows an anastomosing system of irregularly shaped elements. X 24,600.