AqtiQCulture and Fisheries Management 1992, 23, 367-372 Migration of Atlantic salmon, Salmo salar L., parr through a Norwegian fjord: potential infection path of Gyrodactylus salaris R. A. LUND & T. G. HEGGBERGETTVorwg/an Institute for Nature Research, Trondheim, Norway Abstract. Hatchery-reared Atlantic salmon, S(j/mo.«i/arL., parr{0+) ranging in length from 74 to 95 mm were sampled in a stream with no hatchery in its drainage area. Nofishhad previously been released in this stream. The parr, therefore, had migrated during a period when the salinity of the fjord was between 20 and 30 5%o, a minimum distance of 2 7km through the fjord, which was the distance to the nearest hatchery. This is the first observation of pre-smolt migration between streams via a fjord. Earlier, the salmon in the river had been infected by the monogean Gyrodactylus salaris. Probably, this infection had been spread by parr migrating from a neighbouring stream through the fjord. Parasite dispersion coincided with a period of low fjordwater salinity (I2-l4-5%o) and sea temperatures ranging from 8-2 to Xd-S^C, which rs within the suggested tolerance level of G. salaris survival on salmon parr. Introduction Knowledge of the migration of pre-smolt salmon between rivers in coastal areas is limited. During their pre-smolt period salmon parr are normally stationary and migrate within limited areas (Kalleberg 1958; Keenleyside & Yamamoto 1962; Saunders & Gee 1964; Hesthagen 1988, 1990). In central Norway, salmon leave the river as 3- to 4-year-old smolts (Lund & Hansen 1987). During the last decade escapes of cultured fish from fish farms in Norway has increased (Lund, 0k!and & Hansen 1991). However, the escapes of pre-smolt salmon from hatcheries has been less well documented. The present paper investigates the possibility and consequences of escaped pre-smolt salmon migrating from a hatchery to a neighbouring river. The monogean Gyrodactylus salaris has been observed in 32 Norwegian streams (Johnsen, Jensen & Sivertsen 1989) and a serious mortality of salmon has followed in these rivers. Between-river migration of pre-smolt salmon during periods of low salinity might explain the presence of G. salaris in some rivers. Study area The stream Fsttenelva, which is located in mid-Norway, has a catchment area of 12-2kni^, Correspondence; Roar A. Lund, Norwegian Institute for Nature Research, Tungasletta 2, N-7004 Trondheim, Norway. 367
Transcript
AqtiQCulture and Fisheries Management 1992, 23, 367-372
Migration of Atlantic salmon, Salmo salar L., parrthrough a Norwegian fjord: potential infection path ofGyrodactylus salaris
R. A. LUND & T. G. HEGGBERGETTVorwg/an Institute for Nature Research,Trondheim, Norway
Abstract. Hatchery-reared Atlantic salmon, S(j/mo.«i/arL., parr{0+) ranging in length from 74to 95 mm were sampled in a stream with no hatchery in its drainage area. No fish had previouslybeen released in this stream. The parr, therefore, had migrated during a period when thesalinity of the fjord was between 20 and 30 5%o, a minimum distance of 2 7km through thefjord, which was the distance to the nearest hatchery. This is the first observation of pre-smoltmigration between streams via a fjord. Earlier, the salmon in the river had been infected by themonogean Gyrodactylus salaris. Probably, this infection had been spread by parr migratingfrom a neighbouring stream through the fjord. Parasite dispersion coincided with a period oflow fjordwater salinity (I2-l4-5%o) and sea temperatures ranging from 8-2 to Xd-S^C, which rswithin the suggested tolerance level of G. salaris survival on salmon parr.
Introduction
Knowledge of the migration of pre-smolt salmon between rivers in coastal areas is limited.During their pre-smolt period salmon parr are normally stationary and migrate within limitedareas (Kalleberg 1958; Keenleyside & Yamamoto 1962; Saunders & Gee 1964; Hesthagen1988, 1990). In central Norway, salmon leave the river as 3- to 4-year-old smolts (Lund &Hansen 1987).
During the last decade escapes of cultured fish from fish farms in Norway has increased(Lund, 0k!and & Hansen 1991). However, the escapes of pre-smolt salmon from hatcherieshas been less well documented.
The present paper investigates the possibility and consequences of escaped pre-smoltsalmon migrating from a hatchery to a neighbouring river. The monogean Gyrodactylussalaris has been observed in 32 Norwegian streams (Johnsen, Jensen & Sivertsen 1989) and aserious mortality of salmon has followed in these rivers. Between-river migration ofpre-smolt salmon during periods of low salinity might explain the presence of G. salaris insome rivers.
Study area
The stream Fsttenelva, which is located in mid-Norway, has a catchment area of 12-2kni^,
Correspondence; Roar A. Lund, Norwegian Institute for Nature Research, Tungasletta 2, N-7004 Trondheim,Norway.
367
368 R. A. Lund &. T. G. Heggberget
and flows into Trondheimsfjorden (Fig. 1). The stretch which is accessible to upstreammigrating fish is about 8km long. Mean water velocity in this section is 0-3-0-4ni/s. The fishfauna is dominated by brown trout, Salmo trutta L., which are mostly migratory. Salmon, 5.salar L., spawn infrequently in the stream.
A hatchery with a production capacity of 300000 salmon smolts per year is situated on theneighbouring stream, Langsteinelva, which is about 2-7km from the outlet of the Faettenelva.Another hatchery is located about 13km from the Faettenelva (Fig. 1).
Methods and results
Because of infestation by G. salaris, the Fsettenelva was treated with rotenone on 2September 1988. The entire parr population was killed by this. No fish were observed whenthe entire Fattenelva was electrofished on 18 October 1988. However, 6 weeks later (2December) seven salmon parr measuring from 74 to 95 mm were caught by electrofishing inthe iower 800m of the stream, suggesting immigration from the sea.
AU the parr had obvious signs of being reared in a hatchery (Lund, Hansen & Jarvi 1989).Six of them had two or more distorted fins or rounded tail lobes. The remaining parr had onedistorted fin. The fins of all the parr were significantly shorter than the corresponding wildparr fins (Lund et al. 1989). The reared parr also showed a greenish body colour, which isunusual on wild parr, but rather common on hatchery-reared parr. Scale reading indicatedthat four parr were yearlings (0+), and the remaining three were one year old (1 + ).However, scale reading can easily overestimate the age of reared fish because the frequencyof false winter bands is high (Lund & Hansen 1991). All the data, including the fish lengths(Table 1), indicate that all the parr were most probably yearlings.
Discussion
No hatchery is situated in the catchment of the FaetteneWa, and fish have never been releasedin that stream. Consequently, the hatchery-reared parr have immigrated from the fjord. Thelengths and weights (74-95 mm, 4 5-9-5 g) of all the parr were too small to have escaped froma fish farm in the fjord. The external morphology and the scale features of the seven parr
Table 1. Individual characteristics of the salmon parr. TL = total length. CF = condition factor, M = male, F =female, LP = left pectoral fin. RP = right pectoral fin. D = dorsal fin, UT = upper tail lobe, LT = lower tail lobe
Number
1234S67
(mm)
80748580849495
(g)
6 04-57-46-17-08-99-5
CF
117M l1-201-191-181-07M l
Sex
MMFMFMM
Age
0+1 +1 +1 +0+0+0+
LP
159
1314141615
Fin
RP
141414139
1716
length
D
109
U9
105
11
(mm)
UT
14131412141616
LT
141215
n141616
Potential infection path ofG. salaris 369
2 KM
Langsteinelva" • . —
1
Hatchery
Figure 1. Location of the streams examined and the salinity and temperature measurement sites ( • ) .
suggest that these fish have been reared in a hatchery. Therefore, the parr must havemigrated at least 2-7 km through the fjord, which is the shortest distance by sea to the nearesthatchery. An alternative migration distance is 13km to the next hatchery (Fig. 1). Otheralternative origins for the parr are less probable as the distance to other hatcheries is morethan 30km in a fjord area with low freshwater supply (35%o salinity). The streams associatedwith the two nearest hatcheries were examined by electrofishing on 28 September 1988. Highdensities of escaped hatchery-reared parr were observed in both streams.
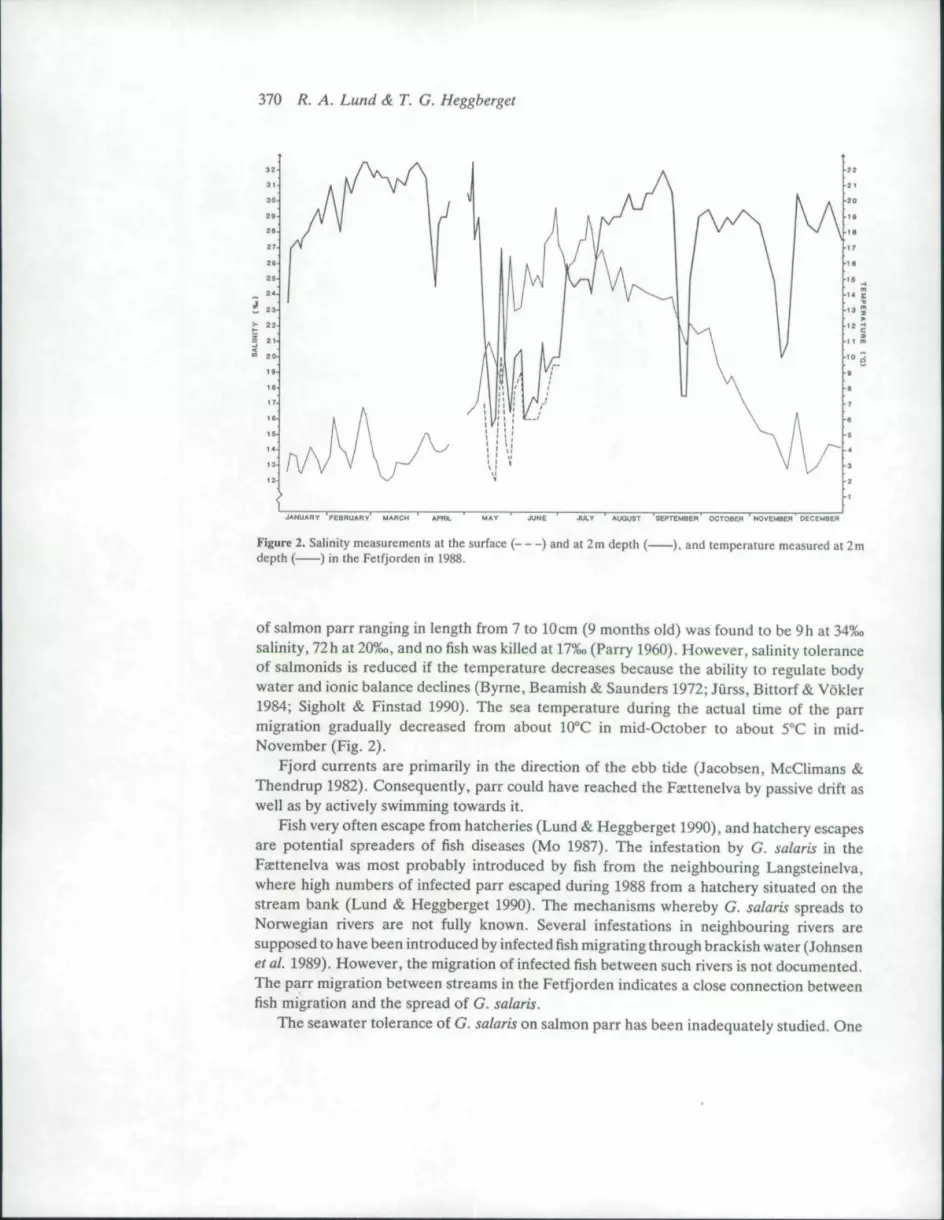
The Feettenetva is smalt, and is easily examined by electrofishing. The stream waselectrofished just before the rotenone treatment, and on 18 October 1988. No salmon parr ofthe actual size of the reared fish, or with morphological characters indicating reared origin,were observed during these fishings. The upstream parr migration, therefore, took placebetween 18 October and 2 December. Salinity recorded at 2 m depth in the fjord basin {Fig. 1)varied between 200 and 30-5% (Fig. 2). However, the salinity in Fetfjorden is usually lowerat the sea surface (Fig. 2).
Although the actual parr migration route is unknown, the fjord salinity would not havebeen a significant barrier. At the experimental temperatures of 10-12°C, mean survival time
370 R. A. Lund & T. G. Heggberget
AUGUST SEPTEMBER OCTOBER NOVEMBEfl OEC EMBER
Figure 2. Salinity measurements at the surface ( ) and at 2m depth ( ), and temperature measured at 2mdepth ( ) in the Fetfjorden in 1988.
of salmon parr ranging in length from 7 to 10cm (9 months old) was found to be 9 h at 34%osalinity, 72h at 20%o, and no fish was killed at 17%o (Parry 1960). However, saHnity toleranceof salmonids is reduced if the temperature decreases because the ability to regulate bodywater and ionic balance declines (Byrne, Beamish & Saunders 1972; Jurss, Bittorf & Vokler1984; Sigholt & Finstad 1990). The sea temperature during the actual time of the parrmigration gradually decreased from about lOX in mid-October to about 5°C in mid-November (Fig. 2).
Fjord currents are primarily in the direction of the ebb tide (Jacobsen, McClimans &Thendrup 1982). Consequently, parr could have reached the Faettenelva by passive drift aswell as by actively swimming towards it.
Fish very often escape from hatcheries (Lund & Heggberget 1990), and hatchery escapesare potential spreaders of fish diseases (Mo 1987). The infestation by G. salaris in theFasttenelva was most probably introduced by fish from the neighbouring Langsteinelva,where high numbers of infected parr escaped during 1988 from a hatchery situated on thestream bank (Lund & Heggberget 1990). The mechanisms whereby G. salaris spreads toNorwegian rivers are not fully known. Several infestations in neighbouring rivers aresupposed to have been introduced by infected fish migrating through brackish water (Johnsenet al 1989). However, the migration of infected fish between such rivers is not documented.The parr migration between streams in the Fetfjorden indicates a close connection betweenfish migration and the spread of G. salaris.
The seawater tolerance of G. salaris on salmon parr has been inadequately studied. One
Potential infection path ofG. salaris 371
study has shown that parasites survived on salmon parr at 8%o salinity at a water temperatureof 10°C (Mo 1986), while another concluded that infected fish had to be kept in 15%o salinewater for 12 h to get rid ofthe parasite (Gyrodactylusprosjektet 1981). In 1988, the salinity inthe Fetfjorden was lowest in the last 2 weeks of May, varying from 12 to 14-5%o (Fig. 2) whilethe sea surface temperature varied from 8 2 to 16-2°C.
The observation that salmon parr migrate between streams in the Fetfjorden and theinfestation of G. salaris in these streams supports the hypothesis that G. salaris has beenspread to several Norwegian rivers by infected fish migrating from neighbouring rivers.Recently, furunculosis (Aeromonas salmonicida) has been observed in several Norwegianstreams. The present observations indicate that diseases, for instance furunculosis, mightalso be spread from hatcheries to neighbouring streams or between streams by fjordmigration of pre-smolt salmon.
References
Byrne J.M., Beamish F.W.H. & Saunders R.L. (1972) InRuence of salinity, temperature, and exercise on plasmaosmolality and ionic concentration in Atlantic salmon (Salmo salar). Journal of ihe Fisheries Research Board ofCanada 29, 1217-1220.
Gyrodactylusprosjektet (1981) Rapport fra Cyrodactylusutvalget over virksomheten I 1980 og program forvirksomheten i I981: 1-59. [In Norwegian.]
Heslhagen T. (1988) Movements of brown troul, Salmo trutta, and juvenile Atlantic salmon, Sfl/mo salar, in a coastalstream in northern Norway. Journal of Fish Biology 32, 639--653.
Hesthagen T. (1990) Home range of juvenile Atlantic salmon, Salmo salar, and brown trout, Salmo trutla. in aNorwegian stream. Freshwater Biology 24, 63-67.
Jacobsen P., McClimans T.A. & Thendrup A. (1982) Vurdering av rensekrav for sjoresipienter. Del 1:Dominerende fysiske prosesser i fjorder og kystfarvann. Norsk. Hydroteknisk Laboratorium, rapport nr.2-83033: 1-76. [In Norwegian.]
Johnsen B.O., Jensen A.J. & Sivertsen B. (1989) Extermination of G^rorfac/^'/iuja/aru-infected Atlantic salmon,Salmo salar, by rctenon treatment in the river Vikja. western Norway. Fauna norvegica Serie A 10, 39-43.
Jurss K., Bittorf T. & Vokler T. (1984) Biochemical investigations on salinity and temperature acclimation oftherainbow trout, Salmo gairdneri (Richardson). Zoologische Jahrbiicher Abieilung fiir Allgemeine Zoologieund Phy.iiologie der Ttere 88, 67-81.
Kalleberg H. (1958) Observation in a stream tank of territoriality and competition in juvenile salmon and trout{Satmo salar L. and 5. trutla L.). Report ofthe Institute of Freshwater Research, Drottningholm 39, 55-59.
Keenleyside M.H.A. & Yamamoto FT. (1962) Territorial behaviour of juvenile Atlantic salmon {Salmo salaris.).Behaviour \9, t39-169.
Lund R.A. & Hansen L.P. (1987) Biological characteristics of Atlantic salmon from coastal fisheries at Kval0ya andKolgrov, western Norway. ICES North Atlantic Salmon Working Group: 1-9
Lund R.A. & Hansen L.P. (1991) Identification of wild and reared Atlantic salmon (Salmo salar) using scalecharacters. Aquaculture and Fisheries Management 22, 499-508.
Lund R.A., Hansen L.P. & Jarvi T. (1989) Identifisering av oppdrettslaks og vilMaks ved ytre morfologi,finnestflrrelse og skjellkarakterer. (Identification of reared and wild salmon by external morphology, size offins and scale characteristics.) NINA Forskningsrapport 1, 1-54. [In Norwegian with English summary.]
Lund R.A. & Heggberget T.G. (1990) Fjordvandring av laksunger, Salmo salar L.; Mulig spredningsvei forGyrodactytus salaris. (Migration of Atlantic salmon parr, Salmo satar L., through a Norwegian fjord. -Potential infection path of Gyrodactytus salaris.} NINA Forskningsrapport S, 1-10. [In Norwegian with Englishabstract.]
Lund R.A., 0kland F. & Hansen L.P. (I99I) Farmed Atlantic salmon (Salmon salar) in fisheries and rivers inNorway. Aquaculture 9S, 143-150.
Mo T.A. (1986) Virkning av rotenon pk laks og lakseparasitten Gyrodactylus salaris. Laboratoriefors0k.Gyrodactylusunders0kelsene ved Zoologisk Museum, Universitetet i Oslo, Rapport nr. 1 1986: 1-16 [InNorwegian.]
372 R. A. Lund & T. G. Heggberget
Mo T.A. (1987) Taksonomiske og biologiske unders0kelser. Virksomheten i 1986 og forslag til virksomhet i 1987.Cyrodactylusunders0kelsene ved Zoologisk Museum, Universitetet i Oslo, Rapport nr. 2; t-70. [In Norwegian.]
Parry G. (1960) The development of the salinity tolerance in the salmon, Satmo satar and some related species.Journal of Experimental Biology 37, 425—434.
Saunders R.L. & Gee J.H. (1964) Movements of young Atlantic salmon in a smali simm. Journat of the FisheriesResearch Board of Canada 21, 27-36.
Sigholt T. & Finstad B. (1990) Effect of low temperature on seawatcr tolerance in Atlantic salmon (Salmo salar)smolts. Aquacutture 84,167-172.