Dual protection of hydroxytyrosol, an olive oilpolyphenol, against oxidative damage inPC12 cells†
Shoujiao Peng, Baoxin Zhang, Juan Yao, Dongzhu Duan and Jianguo Fang*
Hydroxytyrosol (3,4-dihydroxyphenylethanol, HT), a major polyphenol in olive oils, has received increasing
attention due to its multiple pharmacological activities. However, it is not well understood how HT works
on the neuronal system. We report herein that HT efficiently scavenges free radicals in vitro and displays
cytoprotection against oxidative stress-induced damage in PC12 cells. HT completely protects the cells
from hydrogen peroxide-induced death and rescues the cells from 6-hydroxydopamine-induced
damage. Mechanistic studies reveal that Nrf2 is a prerequisite for the neuroprotection of HT as knocking
down Nrf2 eliminated this action. HT, via activation of the Keap1-Nrf2 pathway, elevates a panel of cyto-
protective enzymes, including glutamate–cysteine ligase, HO-1, NQO1 and thioredoxin reductase. Our
study reveals that HT provides dual neuroprotection and cellular antioxidant defense as both a free radical
scavenger and Nrf2 activator, suggesting the potential pharmaceutical usage of HT for the treatment of
neurodegenerative disorders.
Introduction
Reactive oxygen species (ROS) are constantly produced duringnormal cellular metabolism and function as messenger mole-cules in diverse signaling pathways that are involved in celldifferentiation, proliferation, and death.1 However, uponenvironmental stress or cellular damage, cells cannot readilydetoxify ROS and may thereby suffer oxidative stress. Exo-genous antioxidants and cellular endogenous antioxidantsystem are crucial for counteracting ROS-mediated oxidativestress. A well elucidated universal pathway for inducing in-trinsic antioxidant defense involves transcriptional regulationthrough the activation of the antioxidant-responsive element(ARE).2–4 Under basal conditions, the cytosolic regulatoryprotein, Kelch-like ECH-associated protein 1 (Keap1), bindstightly to Nrf2, retaining it in the cytoplasm and targeting it toproteasome degradation.3 The association of Keap1 with Nrf2critically relies on the functional cysteine residue(s) in theprotein Keap1. Substances that could modify these cysteineresidue(s) in the adaptor protein Keap1 would cause therelease of Nrf2 from the inhibitory partner Keap1 and facilitate
its translocation into the nucleus, and thus initiate thetranscription of cytoprotective genes.
The progressive degeneration of a subset of neurons is thepathologic hallmark of adult-onset neurodegenerative dis-orders. As neuronal cells are particularly vulnerable to oxi-dative stress and have limited replenishment during thehuman lifespan, increasing experimental evidence has indi-cated that oxidative stress is a causal, or at least an ancillary,factor in the neuropathology of several adult neurodegenera-tive disorders such as Alzheimer’s disease (AD) and Parkin-son’s disease (PD).5,6 Besides the neutralization of ROS byexogenous small molecule antioxidants, the prevention of oxi-dative stress can be better achieved by the induction ofendogenous cytoprotective systems since their actions aremore sustained and are amplified by transcription-mediatedsignaling cascades. In this sense, activation of the Nrf2-AREpathway is a promising therapeutic approach against neuro-degenerative diseases.4,7 Thus, the past years have witnessedexpanding endeavors that seek to identify naturally occurringand synthetic small molecule activators of the Nrf2-AREpathway as potential neuroprotective agents.8
Hydroxytyrosol (3,4-dihydroxyphenylethanol, HT, Fig. 1A),a naturally occurring polyphenol in table olives and otherdietary plants,9,10 is present in a particularly high concen-tration either in a free or esterified form in extra virginolive oils, wherein it accounts for upto 70%–80% of the totalphenolic fractions.9 The abundance of HT and its esters inolive oils, with concentrations of up to hundred milligrams per
†Electronic supplementary information (ESI) available. See DOI: 10.1039/c5fo00097a
State Key Laboratory of Applied Organic Chemistry and College of Chemistry and
Chemical Engineering, Lanzhou, Gansu 730000, China. E-mail: [email protected];
liter,11 has been suggested to be linked to the health benefitsof the Mediterranean diet.12,13 HT has received increasingattention due to its multiple pharmacological activities suchas antioxidant activity,10,14 cancer cell growth inhibition andapoptosis induction,15,16 and cardiovascular protection.17,18
However, investigations into the neuroprotection of HT arequite limited,19–21 and the underlying mechanism has notbeen fully clarified. In this study, we determined the anti-oxidant activity of HT in vitro and employed the neuron-like ratpheochromocytoma cell line, PC12 cells, to assess the mole-cular mechanisms responsible for the neuroprotective effect ofHT. HT efficiently scavenges free radicals and protects bio-molecules from free radical-induced oxidative damage. Impor-tantly, HT activates the Nrf2-ARE signaling pathway, leading tothe upregulation of phase II enzymes and protection of PC12cells from hydrogen peroxide (H2O2)- or 6-hydroxydopamine(6-OHDA)-induced neurotoxicity. Our study demonstrates thatHT is a potent antioxidant in vitro as well as a novel smallmolecule activator of the Nrf2-ARE pathway in PC12 cells,suggesting that HT might be an encouraging candidate for theprevention of neurodegeneration. Targeting the Nrf2-ARE sig-naling pathway by HT thus provides the molecular mechanismunderlying the neuroprotective effects of HT and sheds lightinto the biological action of HT in vivo.
Results and discussionSynthesis of HT
HT was prepared via a straightforward synthetic route by fol-lowing the procedure described in the literature with minor
modifications.22 Briefly, the esterification of 3,4-dimethoxy-phenylacetic acid gave 3,4-dimethoxyphenylacetic acid methylester, which was further treated with boron tribromide to yield3,4-dihydroxyphenylacetic acid methyl ester. Lithium aluminiumhydride was applied to convert the ester to the desired product.HT was fully characterized by 1HNMR, 13CNMR, and ESI-MS,and the data are consistent with those in the literature.22
Elimination of free radicals in vitro
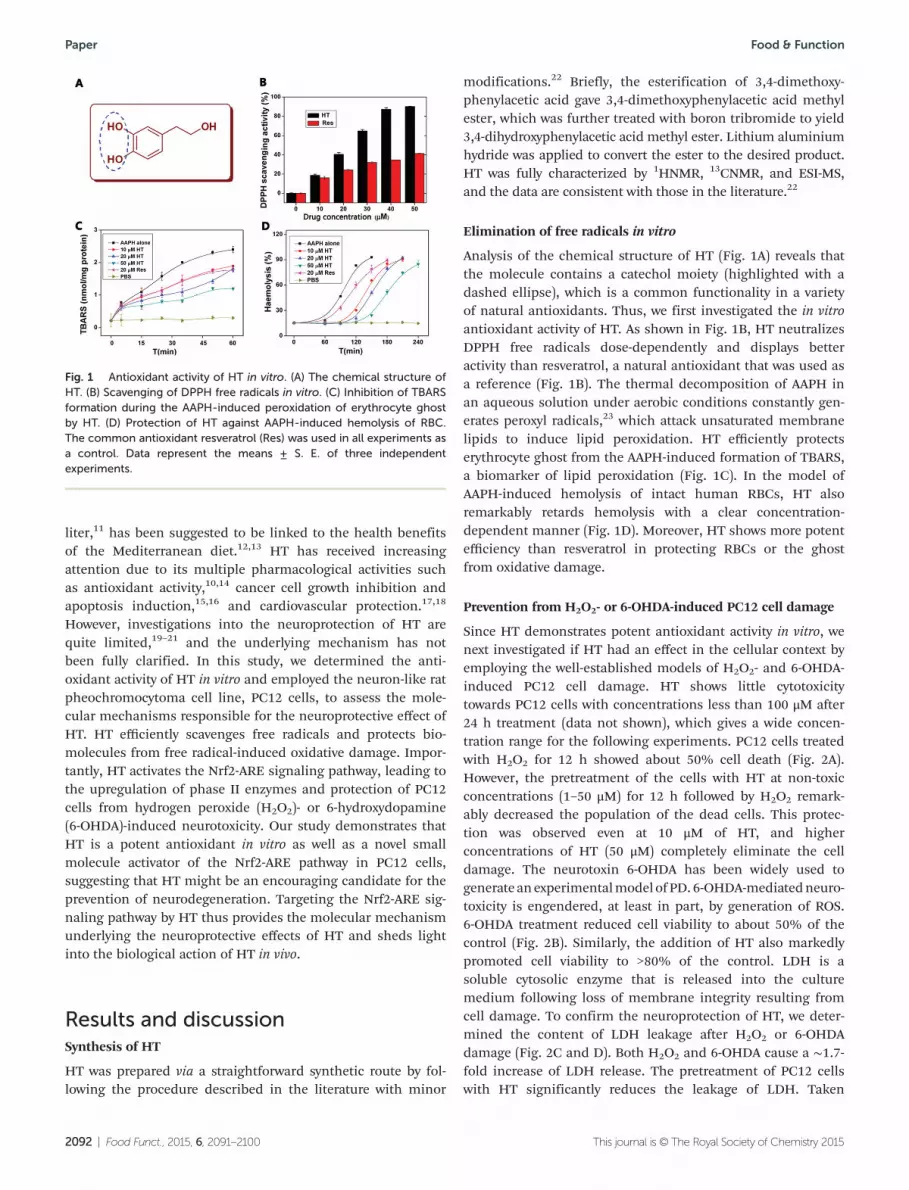
Analysis of the chemical structure of HT (Fig. 1A) reveals thatthe molecule contains a catechol moiety (highlighted with adashed ellipse), which is a common functionality in a varietyof natural antioxidants. Thus, we first investigated the in vitroantioxidant activity of HT. As shown in Fig. 1B, HT neutralizesDPPH free radicals dose-dependently and displays betteractivity than resveratrol, a natural antioxidant that was used asa reference (Fig. 1B). The thermal decomposition of AAPH inan aqueous solution under aerobic conditions constantly gen-erates peroxyl radicals,23 which attack unsaturated membranelipids to induce lipid peroxidation. HT efficiently protectserythrocyte ghost from the AAPH-induced formation of TBARS,a biomarker of lipid peroxidation (Fig. 1C). In the model ofAAPH-induced hemolysis of intact human RBCs, HT alsoremarkably retards hemolysis with a clear concentration-dependent manner (Fig. 1D). Moreover, HT shows more potentefficiency than resveratrol in protecting RBCs or the ghostfrom oxidative damage.
Prevention from H2O2- or 6-OHDA-induced PC12 cell damage
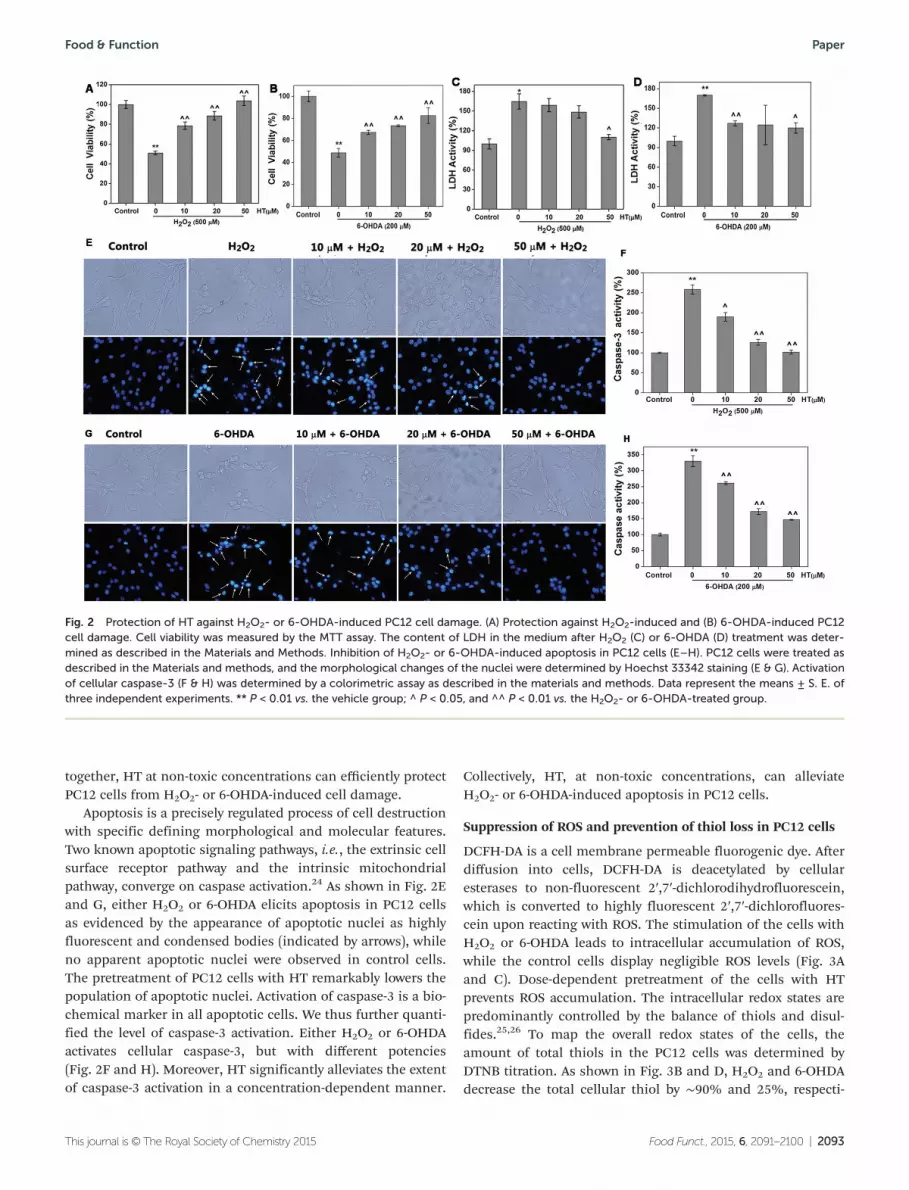
Since HT demonstrates potent antioxidant activity in vitro, wenext investigated if HT had an effect in the cellular context byemploying the well-established models of H2O2- and 6-OHDA-induced PC12 cell damage. HT shows little cytotoxicitytowards PC12 cells with concentrations less than 100 μM after24 h treatment (data not shown), which gives a wide concen-tration range for the following experiments. PC12 cells treatedwith H2O2 for 12 h showed about 50% cell death (Fig. 2A).However, the pretreatment of the cells with HT at non-toxicconcentrations (1–50 μM) for 12 h followed by H2O2 remark-ably decreased the population of the dead cells. This protec-tion was observed even at 10 μM of HT, and higherconcentrations of HT (50 μM) completely eliminate the celldamage. The neurotoxin 6-OHDA has been widely used togenerate anexperimentalmodel ofPD. 6-OHDA-mediatedneuro-toxicity is engendered, at least in part, by generation of ROS.6-OHDA treatment reduced cell viability to about 50% of thecontrol (Fig. 2B). Similarly, the addition of HT also markedlypromoted cell viability to >80% of the control. LDH is asoluble cytosolic enzyme that is released into the culturemedium following loss of membrane integrity resulting fromcell damage. To confirm the neuroprotection of HT, we deter-mined the content of LDH leakage after H2O2 or 6-OHDAdamage (Fig. 2C and D). Both H2O2 and 6-OHDA cause a ∼1.7-fold increase of LDH release. The pretreatment of PC12 cellswith HT significantly reduces the leakage of LDH. Taken
Fig. 1 Antioxidant activity of HT in vitro. (A) The chemical structure ofHT. (B) Scavenging of DPPH free radicals in vitro. (C) Inhibition of TBARSformation during the AAPH-induced peroxidation of erythrocyte ghostby HT. (D) Protection of HT against AAPH-induced hemolysis of RBC.The common antioxidant resveratrol (Res) was used in all experiments asa control. Data represent the means ± S. E. of three independentexperiments.
together, HT at non-toxic concentrations can efficiently protectPC12 cells from H2O2- or 6-OHDA-induced cell damage.
Apoptosis is a precisely regulated process of cell destructionwith specific defining morphological and molecular features.Two known apoptotic signaling pathways, i.e., the extrinsic cellsurface receptor pathway and the intrinsic mitochondrialpathway, converge on caspase activation.24 As shown in Fig. 2Eand G, either H2O2 or 6-OHDA elicits apoptosis in PC12 cellsas evidenced by the appearance of apoptotic nuclei as highlyfluorescent and condensed bodies (indicated by arrows), whileno apparent apoptotic nuclei were observed in control cells.The pretreatment of PC12 cells with HT remarkably lowers thepopulation of apoptotic nuclei. Activation of caspase-3 is a bio-chemical marker in all apoptotic cells. We thus further quanti-fied the level of caspase-3 activation. Either H2O2 or 6-OHDAactivates cellular caspase-3, but with different potencies(Fig. 2F and H). Moreover, HT significantly alleviates the extentof caspase-3 activation in a concentration-dependent manner.
Collectively, HT, at non-toxic concentrations, can alleviateH2O2- or 6-OHDA-induced apoptosis in PC12 cells.
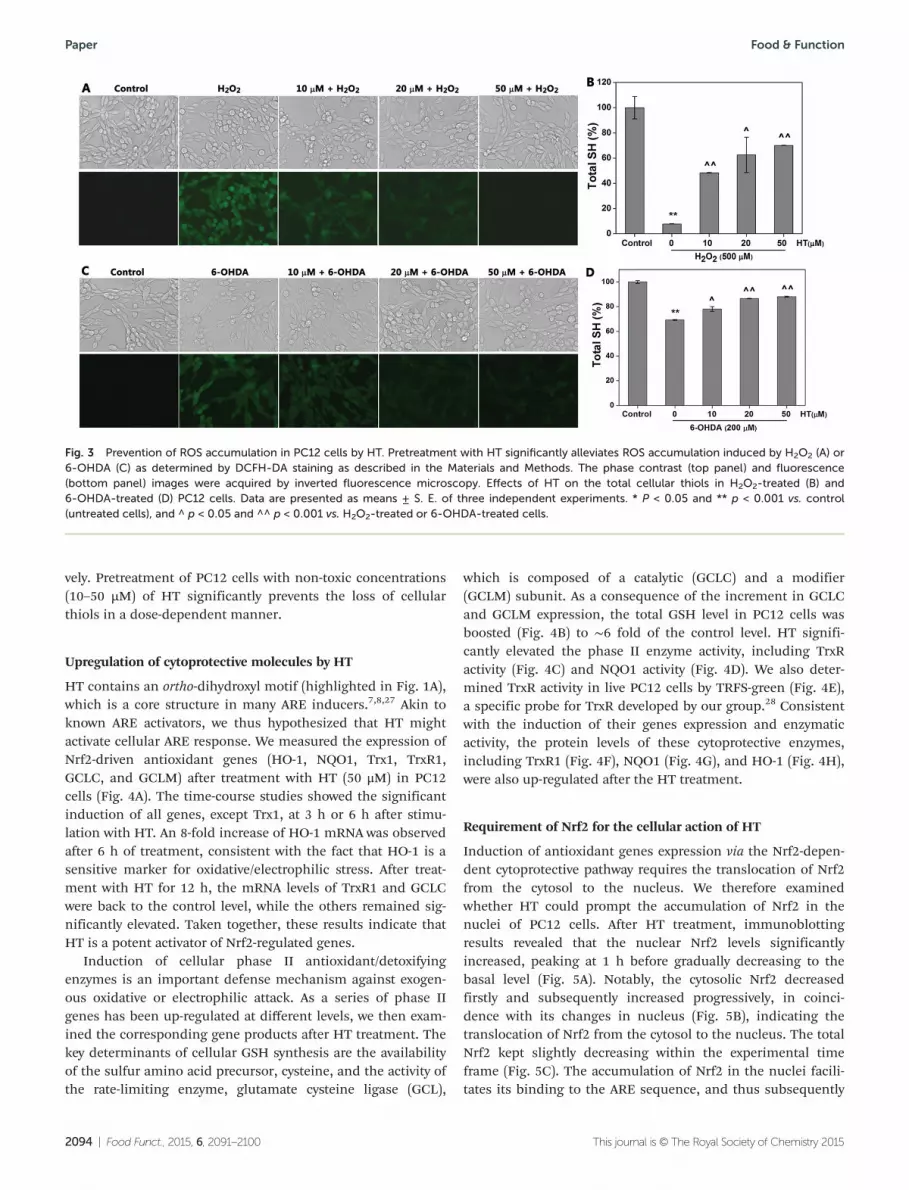
Suppression of ROS and prevention of thiol loss in PC12 cells
DCFH-DA is a cell membrane permeable fluorogenic dye. Afterdiffusion into cells, DCFH-DA is deacetylated by cellularesterases to non-fluorescent 2′,7′-dichlorodihydrofluorescein,which is converted to highly fluorescent 2′,7′-dichlorofluores-cein upon reacting with ROS. The stimulation of the cells withH2O2 or 6-OHDA leads to intracellular accumulation of ROS,while the control cells display negligible ROS levels (Fig. 3Aand C). Dose-dependent pretreatment of the cells with HTprevents ROS accumulation. The intracellular redox states arepredominantly controlled by the balance of thiols and disul-fides.25,26 To map the overall redox states of the cells, theamount of total thiols in the PC12 cells was determined byDTNB titration. As shown in Fig. 3B and D, H2O2 and 6-OHDAdecrease the total cellular thiol by ∼90% and 25%, respecti-
Fig. 2 Protection of HT against H2O2- or 6-OHDA-induced PC12 cell damage. (A) Protection against H2O2-induced and (B) 6-OHDA-induced PC12cell damage. Cell viability was measured by the MTT assay. The content of LDH in the medium after H2O2 (C) or 6-OHDA (D) treatment was deter-mined as described in the Materials and Methods. Inhibition of H2O2- or 6-OHDA-induced apoptosis in PC12 cells (E–H). PC12 cells were treated asdescribed in the Materials and methods, and the morphological changes of the nuclei were determined by Hoechst 33342 staining (E & G). Activationof cellular caspase-3 (F & H) was determined by a colorimetric assay as described in the materials and methods. Data represent the means ± S. E. ofthree independent experiments. ** P < 0.01 vs. the vehicle group; ^ P < 0.05, and ^^ P < 0.01 vs. the H2O2- or 6-OHDA-treated group.
vely. Pretreatment of PC12 cells with non-toxic concentrations(10–50 µM) of HT significantly prevents the loss of cellularthiols in a dose-dependent manner.
Upregulation of cytoprotective molecules by HT
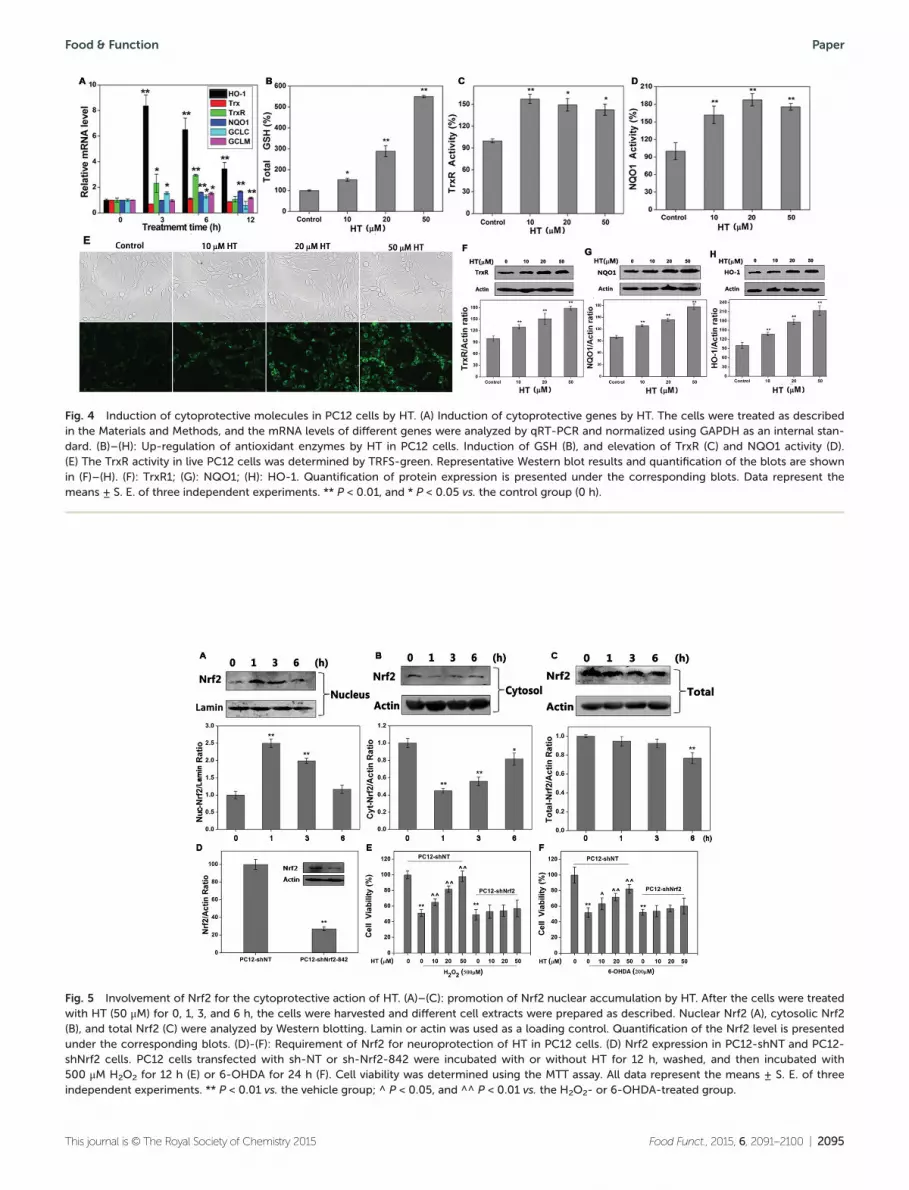
HT contains an ortho-dihydroxyl motif (highlighted in Fig. 1A),which is a core structure in many ARE inducers.7,8,27 Akin toknown ARE activators, we thus hypothesized that HT mightactivate cellular ARE response. We measured the expression ofNrf2-driven antioxidant genes (HO-1, NQO1, Trx1, TrxR1,GCLC, and GCLM) after treatment with HT (50 μM) in PC12cells (Fig. 4A). The time-course studies showed the significantinduction of all genes, except Trx1, at 3 h or 6 h after stimu-lation with HT. An 8-fold increase of HO-1 mRNA was observedafter 6 h of treatment, consistent with the fact that HO-1 is asensitive marker for oxidative/electrophilic stress. After treat-ment with HT for 12 h, the mRNA levels of TrxR1 and GCLCwere back to the control level, while the others remained sig-nificantly elevated. Taken together, these results indicate thatHT is a potent activator of Nrf2-regulated genes.
Induction of cellular phase II antioxidant/detoxifyingenzymes is an important defense mechanism against exogen-ous oxidative or electrophilic attack. As a series of phase IIgenes has been up-regulated at different levels, we then exam-ined the corresponding gene products after HT treatment. Thekey determinants of cellular GSH synthesis are the availabilityof the sulfur amino acid precursor, cysteine, and the activity ofthe rate-limiting enzyme, glutamate cysteine ligase (GCL),
which is composed of a catalytic (GCLC) and a modifier(GCLM) subunit. As a consequence of the increment in GCLCand GCLM expression, the total GSH level in PC12 cells wasboosted (Fig. 4B) to ∼6 fold of the control level. HT signifi-cantly elevated the phase II enzyme activity, including TrxRactivity (Fig. 4C) and NQO1 activity (Fig. 4D). We also deter-mined TrxR activity in live PC12 cells by TRFS-green (Fig. 4E),a specific probe for TrxR developed by our group.28 Consistentwith the induction of their genes expression and enzymaticactivity, the protein levels of these cytoprotective enzymes,including TrxR1 (Fig. 4F), NQO1 (Fig. 4G), and HO-1 (Fig. 4H),were also up-regulated after the HT treatment.
Requirement of Nrf2 for the cellular action of HT
Induction of antioxidant genes expression via the Nrf2-depen-dent cytoprotective pathway requires the translocation of Nrf2from the cytosol to the nucleus. We therefore examinedwhether HT could prompt the accumulation of Nrf2 in thenuclei of PC12 cells. After HT treatment, immunoblottingresults revealed that the nuclear Nrf2 levels significantlyincreased, peaking at 1 h before gradually decreasing to thebasal level (Fig. 5A). Notably, the cytosolic Nrf2 decreasedfirstly and subsequently increased progressively, in coinci-dence with its changes in nucleus (Fig. 5B), indicating thetranslocation of Nrf2 from the cytosol to the nucleus. The totalNrf2 kept slightly decreasing within the experimental timeframe (Fig. 5C). The accumulation of Nrf2 in the nuclei facili-tates its binding to the ARE sequence, and thus subsequently
Fig. 3 Prevention of ROS accumulation in PC12 cells by HT. Pretreatment with HT significantly alleviates ROS accumulation induced by H2O2 (A) or6-OHDA (C) as determined by DCFH-DA staining as described in the Materials and Methods. The phase contrast (top panel) and fluorescence(bottom panel) images were acquired by inverted fluorescence microscopy. Effects of HT on the total cellular thiols in H2O2-treated (B) and6-OHDA-treated (D) PC12 cells. Data are presented as means ± S. E. of three independent experiments. * P < 0.05 and ** p < 0.001 vs. control(untreated cells), and ^ p < 0.05 and ^^ p < 0.001 vs. H2O2-treated or 6-OHDA-treated cells.
Fig. 4 Induction of cytoprotective molecules in PC12 cells by HT. (A) Induction of cytoprotective genes by HT. The cells were treated as describedin the Materials and Methods, and the mRNA levels of different genes were analyzed by qRT-PCR and normalized using GAPDH as an internal stan-dard. (B)–(H): Up-regulation of antioxidant enzymes by HT in PC12 cells. Induction of GSH (B), and elevation of TrxR (C) and NQO1 activity (D).(E) The TrxR activity in live PC12 cells was determined by TRFS-green. Representative Western blot results and quantification of the blots are shownin (F)–(H). (F): TrxR1; (G): NQO1; (H): HO-1. Quantification of protein expression is presented under the corresponding blots. Data represent themeans ± S. E. of three independent experiments. ** P < 0.01, and * P < 0.05 vs. the control group (0 h).
Fig. 5 Involvement of Nrf2 for the cytoprotective action of HT. (A)–(C): promotion of Nrf2 nuclear accumulation by HT. After the cells were treatedwith HT (50 μM) for 0, 1, 3, and 6 h, the cells were harvested and different cell extracts were prepared as described. Nuclear Nrf2 (A), cytosolic Nrf2(B), and total Nrf2 (C) were analyzed by Western blotting. Lamin or actin was used as a loading control. Quantification of the Nrf2 level is presentedunder the corresponding blots. (D)-(F): Requirement of Nrf2 for neuroprotection of HT in PC12 cells. (D) Nrf2 expression in PC12-shNT and PC12-shNrf2 cells. PC12 cells transfected with sh-NT or sh-Nrf2-842 were incubated with or without HT for 12 h, washed, and then incubated with500 μM H2O2 for 12 h (E) or 6-OHDA for 24 h (F). Cell viability was determined using the MTT assay. All data represent the means ± S. E. of threeindependent experiments. ** P < 0.01 vs. the vehicle group; ^ P < 0.05, and ^^ P < 0.01 vs. the H2O2- or 6-OHDA-treated group.
initiates the transcription process to induce phase II geneexpression.
As a set of Nrf2-driven genes and the related gene productswere elevated in PC12 cells after HT treatment, we performedNrf2 knockdown experiments to further clarify whether HTprotects PC12 cells via the activation of Nrf2. The knockdownefficiency of shNrf2 in PC12 cells was validated by Westernblots (Fig. 5D). The expression of Nrf2 protein in thecells transfected with shNrf2 (PC12-shNrf2) was drasticallydecreased to ∼30% of that in those transfected with thecontrol non-targeting plasmid (PC12-shNT). Next, we evaluatedthe protective effect of HT against oxidative stress toward thecells. As shown in Fig. 5E and F, HT displays a similar protec-tive pattern in PC12-shNT cells as that observed in normalPC12 cells (Fig. 2A and B). However, this protection dropsstrikingly in PC12-shNrf2 cells. HT, at low concentrations (10 &20 μM), totally lost its ability to rescue PC12-shNrf2 cells andonly marginal protection was observed at high concentration(50 μM). This marginal protection is possibly due to the directantioxidant activity of HT, as shown in Fig. 1. Our resultsfirmly evidenced that Nrf2 is required for the cytoprotection ofHT against oxidative stress in PC12 cells.
Medicinal chemical properties of HT
The medicinal chemical properties of a molecule could predictwhether it could be a successful central nervous system (CNS)drug. The general criteria for a good CNS drug candidate areas follows: the molecular weight (MW) is less than 450, clog Pis between 0 and 5, and the polar surface area (PSA) is lessthan 70 Å2.29 HT has a MW of 154.2. By determination of thepartition coefficient between n-octanol and water, the clog Pvalue of HT was found to be 1.07. The positive control o-cresolgives a value of 1.95, consistent with the data in the reference(1.98 ± 0.05).49 The PAS value was calculated to be 60.7. Thus,the medicinal chemical properties of HT support it as a goodcandidate as a CNS drug.
Neuronal cells in brains are particularly vulnerable to oxi-dative stress as they possess relatively restricted replenishmentfrom progenitor cells, high levels of oxidizable unsaturatedfatty acids, abundance of redox-active metals (e.g. iron andcopper), high oxygen consumption, and low levels of anti-oxidant defense capacities.30 In line with the above-describedsituation, there has been increasing evidence underpinningoxidative stress as a major cause in a number of neurodegen-erative disorders such as AD and PD.5,6 Clinical treatment ofthese diseases by replacing damaged neurons has beendemonstrated with limited success.31 Therefore, pharmaco-logical interventions aimed at the alleviation of oxidative stressare promising approaches to confer protection to neuronalcells, eventually leading to retardation or blockage of the pro-gression of these neurodegenerative diseases. In this regard,there is a clear rationale for the discovery and development ofnovel small molecules that can induce endogenous antioxidantdefenses as potential therapeutic agents for neurodegenerativediseases.8,32
In our continuing interest in discovering and developingnovel small molecule regulators of the cellular redoxsystem,28,33–37 we demonstrated here that HT, an active con-stituent of olive oils, is a promising neuroprotective agent. Wealso revealed the mechanism underlying the action of HT inPC12 cells. HT is fairly stable in the cell culture medium.HPLC analysis indicates that ∼75% of HT remained in the cellmedium after a 12 h incubation at 37 degree. The recovery ofHT is ∼45% after incubating HT with PC12 cells for 12 h(Fig. S1 in the ESI†). HT acts as both a direct antioxidant forscavenging free radicals in vitro and confers protective effectsagainst oxidative stress-induced PC12 cell damage by activatingthe cellular antioxidant defense systems. The high antioxidantactivity and low cytotoxicity of HT make it an ideal moleculefor further development as a potential therapeutic agent. Likemany phenolic compounds, the direct antioxidant action ofHT depends primarily on its ability to donate its phenolicH-atom to different radicals via a hydrogen abstraction reac-tion, and the ortho-dihydroxyl functionality further enhancesits free radical-scavenging activity.23 It was reported that HTactivates Nrf2 in vascular endothelial cells,18 HepG2 cells38
and retinal pigment epithelial cells.39 A recent study alsosuggested that HT confers neuroprotection via the inhibitionof catechol-O-methyl transferase activity in rat striatal tissue.40
Our results show that HT, via the activation of the Keap1-Nrf2-ARE pathway, up-regulated the expression of HO-1, NQO1,TrxR1, GCLM, and GCLC genes and increased their gene pro-ducts in PC12 cells. Clearly, the induction of these cytoprotec-tive molecules could counteract oxidative stress-mediatedtoxicity. The mechanisms underlying the activation of Nrf2 arenot well understood but may involve the oxidation/alkylationof key thiols in Keap1 and/or phosphorylation of Nrf2.41 Uponexposure to chemical stresses, the Cys-rich protein Keap1 ismodified by forming a covalent adduct or disulfide bondwithin the protein, and it is this Cys-based modification ofKeap1 that allows for dissociation from Nrf2 and subsequentnuclear translocation of Nrf2, wherein it binds to ARE andinitiates the transcription of cytoprotective genes. The obser-vation that HT causes the nuclear accumulation of Nrf2 andthat knocking down Nrf2 attenuates cytoprotection supportsthe involvement of HT in the Nrf2-ARE pathway. Analysis ofthe chemical structure of HT reveals that the molecule con-tains the ortho-dihydroxyl moiety (highlighted in Fig. 1A),which is easily oxidized to ortho-quinone, a thiol-reactiveelectrophilic intermediate that may covalently modify the Cysthiols. Thus, HT may be regarded as a proelectrophile.27 Manynatural or synthetic molecules bearing this structure such ascarnosic acid are known proelectrophiles, which induce phaseII genes.42 Thus, we speculated that the alkylation of certainCys residue(s) by the ortho-quinone form of HT might be themolecular basis for the activation of the Keap1-Nrf2-AREpathway by HT.
Our findings suggest a dual neuroprotective mechanism ofHT, shown schematically in Fig. 6. The lipophilic character(clog P = 1.07) and small size (MW = 154.2 and PSA = 60.7) ofHT allow it to easily pass through the plasma membrane and
get into cells. As a phenolic compound, HT directly counter-acts various ROS. More importantly, HT activates the cellularendogenous defense network by the upregulation of diverseantioxidant molecules such as GSH, NQO1, TrxR1 and HO-1.Keap1 serves as an adaptor for ubiquitin E3 ligase and pro-motes the proteasomal degradation of Nrf2 in quiescent cells.Nrf2 is stabilized when Keap1 is inactivated under oxidative/electrophilic stress conditions. HT, presumably via its quinoneintermediate (dashed square in Fig. 6) resulting from the reac-tion with ROS, may modify the Cys residue(s) on the cytosolicprotein Keap1, and dissociate the Keap1-Nrf2 complex, leadingto the liberation of Nrf2 and enabling it to be translocated intonucleus, wherein it binds to ARE and starts the transcriptionof cytoprotective genes. HT directly neutralizes free radicalsand indirectly induces the endogenous antioxidant defensesystem in cells, thus providing a dual protection against oxi-dative stress-induced neuronal cell damage.
Materials and methodsMaterials
Dulbecco’s modified Eagle’s medium (DMEM), N-acetyl-Asp-Glu-Val-Asp-p-nitroanilide (Ac-DEVD-pNA), dimethyl sulfoxide(DMSO), yeast glutathione reductase (GR), Hoechst 33342 and2′,7′-dichlorfluorescein diacetate (DCFH-DA), 2,2-diphenyl-1-picrylhydrazyl (DPPH), 5,5′-dithiobis-2-nitrobenzoic acid(DTNB), 2,6-dichlorophenol-indophenol (DCPIP), NADH, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT), 2,2′-azobis-2-methyl-propanimidamide dihydrochloride(AAPH), penicillin and streptomycin were obtained fromSigma-Aldrich (St. Louis, USA). NADPH were obtained fromRoche (Mannheim, Germany). Fetal bovine serum (FBS) wasobtained from HyClone. 6-OHDA, hydrogen peroxide, anti-bodies against TrxR1, actin, and Nrf2 were from Santa CruzBiotechnology. Thiobarbituric acid (TBA) and antibodiesagainst HO-1, NQO1 and lamin were from Sangon Biotech(Shanghai, China). The shRNA plasmids targeting the codingregions of the rat Nrf2 gene (shNrf2) and the control non-targeting shRNA (shNT) were purchased from GenePharma Co,Ltd (Shanghai, China). GeneTran III transfection reagent wasobtained from Biomiga (CA, USA). Bovine serum albumin(BSA), butylated hydroxytoluene (BHT) and dithiothreitol
(DTT) were from Beyotime (Nantong, China). HT was dissolvedin DMSO, and the final DMSO concentration in all cell experi-ments was not higher than 0.1% (v/v). All other reagents wereof analytical grade.
Cell culture
The rat adrenal pheochromocytoma cell line, PC12, wasobtained from the Shanghai Institute of Biochemistry and CellBiology, Chinese Academy of Sciences, and was cultured inDMEM supplemented with 10% FBS, 2 mM glutamine, and100 units per ml penicillin/streptomycin and maintained in ahumidified atmosphere of 5% CO2 at 37 °C.
In vitro antioxidant activities
The DPPH free radical scavenging assay, free radical-inducedhemolysis of red blood cells (RBC) and formation of the thio-barbituric acid reactive substances (TBARS) in erythrocyteghosts are described in our previously published pro-cedures.33,43 Varying concentrations of HT were mixed withthe DPPH solution (100 μM) at room temperature for 30 minin the dark. The absorbance of the samples was measuredspectrophotometrically at 517 nm. For the RBC hemolysisassay, 5% suspension of RBC in PBS was incubated under airat 37 °C for 5 min. AAPH (final concentration 50 mM) wasadded to initiate hemolysis. The extent of hemolysis was deter-mined by measuring the absorbance of the supernatant at540 nm. For measurements of TBARS, the erythrocyte ghost(final protein concentration of 0.96 mg ml−1) was incubated at37 °C in 0.1 M PBS. Peroxidation was initiated by the additionof AAPH (final concentration 50 mM). The reaction mixturewas gently shaken at 37 °C and aliquots of the reactionmixture were taken out at specific intervals to determine theformation of TBARS using the extinction coefficient of 1.56 ×105 M−1 cm−1 at 532 nm.
MTT assay and Lactate dehydrogenase (LDH) release assay
To evaluate the cytotoxicity of HT, PC12 cells (1 × 104 cells perwell) were seeded in 96-well plates for 1 day followed by incu-bation with HT for 24 h, after which cell viability was deter-mined by the MTT assay. For the H2O2 or 6-OHDA injurymodel, PC12 cells (1 × 104 cells per well) were plated in a 96-well plate and allowed to adhere for 24 h, and then treatedwith the drug for 12 h. After replacing the medium with freshmedium containing 500 μM H2O2 for 12 h or 200 μM 6-OHDAfor 24 h, cell viability was determined by the MTT assay.37 Forthe LDH release assay, the cells (2 × 105 cells per well) wereplated in 12-well plates. On the following day, the cells wereexposed to varying concentrations of HT for 12 h, followed bythe addition of 500 μM H2O2 for additional 12 h or 200 μM6-OHDA for additional 24 h. The leakage of LDH from the cul-tured cells was quantified by measuring LDH activity in theculture medium according to our published procedures.33,37
Apoptosis assays
Hoechst 33342 staining. PC12 cells (2 × 105 cells per well)were seeded into 12-well plates. After 24 h, the cells were
Fig. 6 A dual neuroprotective mode of HT in PC12 cells.
treated with HT for another 24 h followed by replacing themedium with fresh medium containing 500 μM H2O2 for 12 hor 200 μM 6-OHDA for 24 h. Hoechst 33342 was subsequentlyadded to a final concentration of 5 μg ml−1 to stain the nuclei.The cells were visualized and photographed under a Leicainverted fluorescent microscope. Cells displaying condensedand highly fluorescent nuclei, a characteristic morphology ofcells undergoing apoptosis, were counted as apoptotic cells.
Measurement of caspase-3 activity. For determination ofcaspase-3 activity, PC12 cells (1 × 106 cells per well) wereseeded in 60 mm dishes for 24 h and then treated withdifferent concentrations of HT for another 24 h. After replacingthe medium with fresh medium containing 500 μM H2O2 or200 μM 6-OHDA and culturing for 24 h, the cells were lysedwith RIPA buffer (50 mM Tris-HCl, pH 7.5, 2 mM EDTA, 0.5%deoxycholate, 150 mM NaCl, 1% Triton X-100, 0.1% SDS,1 mM Na3VO4 and 1 mM PMSF). Protein content was quanti-fied using the Bradford procedure. The caspase-3 activity inthe lystae was determined by following the published pro-cedure.44,45 Briefly, a cell extract containing 15 μg of total pro-teins was incubated with the assay mixture (50 mM Hepes, pH7.5, 2 mM EDTA, 5% glycerol, 10 mM DTT, 0.1% CHAPS,0.2 mM Ac-DEVD-pNA) for 5 h at 37 °C in a final volume of100 μl. The absorbance at 405 nm was measured using amicroplate reader. The activity was expressed as the percentageof the control cells without any drug treatment.
Determination of intracellular ROS
For assaying the intracellular accumulation of ROS,45–47 PC12cells (2 × 105 cells per well) were seeded into 12-well plates.After 24 h, the cells were treated with HT for another 24 h fol-lowed by replacing the medium with fresh medium containing500 μM H2O2 for 12 h or 200 μM 6-OHDA for 5 h. After remov-ing the medium, DCFH-DA (10 μM) in fresh FBS-free mediumwas added and the cells were incubated for 30 min at 37 °C ina CO2 incubator. The cells were visualized and photographedby fluorescence microscopy. The appearance of green fluo-rescence indicates the accumulation of ROS in cells.
Assessment of intracellular thiols
PC12 cells were plated in 60 mm dishes at a concentration of8 × 105 cells per well and allowed to adhere for 24 h. The cellswere then treated with HT (0–50 µM) for 12 h. After replacingthe medium with fresh medium containing H2O2 (500 μM) or6-OHDA (200 μM) for 24 h, the cells were collected, washedtwice with PBS, and lysed with RIPA buffer for 30 min on ice.Protein content was quantified using the Bradford procedure.Total thiols were determined by DTNB-titration.45,48 Briefly,10 µl cell lysate was added to cuvettes containing 90 μl of1 mM DTNB in 6 M guanidine hydrochloride, pH 8.0. After5 min at room temperature, the absorbance was read at412 nm. Thiol levels were calculated by a calibration curveusing glutathione (GSH) as the standard.
Real-time reverse transcription-PCR (RT-PCR)
PC12 cells (1 × 106 cells per well) were seeded in 60 mm dishesfor 24 h, and treated with 50 μM HT for 0, 3, 6 and 12 h. TotalRNA was isolated from cells using the RNAiso plus accordingto the manufacture’s protocol (TaKaRa, Dalian, China) andquantified through 260/280 absorbance. Reverse transcriptionwas performed using a Primescript™ RT reagent kit accordingto the manufacture’s protocol (TaKaRa, Dalian, China). ThePCR reactions were performed on a Mx3005P RT-PCR System(Agilent Technologies) using SYBR Green PCR Master Mix(TaKaRa, Dalian, China). Glyceraldehyde-3-phosphate dehydro-genase (GAPDH) was used as an internal control. The targetgene expression was measured and was normalized to theGAPDH expression level. PCR primers specific to each gene areas follows: GAPDH: 5′-cagtgccagcctcgtctcat-3′ and 5′-aggggc-catccacagtcttc-3′; Heme oxygenase 1 (HO-1): 5′-gccctggaagagga-gatagag-3′ and 5′-tagtgctgtgtggctggtgt-3′; NAD(P)H: quinoneoxidoreductase 1 (NQO1): 5′-tcaccactctactttgctccaa-3′ and5′-ttttctgctcctcttgaacctc-3′; thioredoxin 1 (Trx1): 5′-ccttctttca-ttccctctgtgac-3′ and 5′-cccaaccttttgaccctttttat-3′; thioredoxinreductase 1 (TrxR1): 5′-actgctcaatccacaaacagc-3′ and 5′-ccacggt-ctctaagccaatagt-3′; glutamate cysteine ligase catalytic subunit(GCLC): 5′-caaggacaagaacacaccatct-3′ and 5′-cagcactcaaagcca-taacaat-3′; glutamate cysteine ligase modifier (GCLM): 5′-ggca-caggtaaaacccaatagt-3′ and 5′-ttcaatgtcagggatgctttct-3′.
Measurement of total GSH, NQO1 activity and TrxR activity
PC12 cells were seeded in 60 mm dishes for 24 h and thentreated with different concentrations of HT for another 24 h.The cells were harvested and the detailed procedure for deter-mination of total glutathione, NQO1 activity and TrxR activityin PC12 cell lysate were followed as described in our recentlypublished work.33,37 Total glutathione was assayed by the enzy-matic method. The cells were lysed by KPE buffer (0.1%TritonX-100 and 0.6% sulfosalicyclic acid in 0.1 M PBS with5 mM EDTA, pH 7.5). 120 μl of master mixture containing0.33 mg ml−1 DTNB and 1.66 units per ml glutathionereductase was added to each cell lystae (20 μl). β-NADPH (60 μlof 0.66 mg ml−1) was then added quickly and the absorbanceat 412 nm was measured. The total glutathione content in thesamples was determined by comparison with the pre-determined glutathione standard curve. For measuring NQO1,Trx, and TrxR activity, the cells were lysed in RIPA buffer. Totalprotein content was quantified using the Bradford procedure.NQO1 activity was determined spectrophotometrically bymonitoring the reduction of the electron acceptor, 2,6-dichloro-phenol-indophenol (DCPIP), at 600 nm with or withoutNQO1 inhibitor dicoumarol. The dicoumarol-inhibitable partof DCPIP’s reduction was used to calculate NQO1 activity. Todetermine the activity of TrxR, the cell extract was incubatedwith insulin, NADPH, and recombinant Trx for 30 min at37 °C. To determine the activity of Trx, the cell extract wasincubated with insulin, NADPH, and recombinant rat TrxR1for 30 min at 37 °C. The reaction was terminated by addingDTNB in 6 M guanidine hydrochloride. A blank sample con-
taining everything except Trx (for TrxR assay) or TrxR (for Trxassay) was treated in the same manner. The absorbance at412 nm was measured, and the blank value was subtractedfrom the corresponding absorbance value of the sample.
Imaging TrxR activity in live PC12 cells by TRFS-green28,44
PC12 cells were treated with the indicated concentrations ofHT for 24 h, followed by treatment with TRFS-green (10 µM)for additional 4 h. Phase contrast and fluorescence imageswere acquired with a Floid™ cell imaging station (Life Tech-nology). The intensity of the green fluorescence in cells indi-cates the relative cellular TrxR activity.
Preparation of different protein extracts for Western blotanalysis
The preparation of whole cell protein extraction, cytosolicprotein fraction and nuclear protein extracts was conductedaccording to our published procedures.33,37 For the whole cellprotein extraction, PC12 cells were lysed with RIPA. For thecytosolic and nuclear extracts, the cells were rinsed with ice-cold PBS and resolved in 100 μL of buffer A (10 mM Hepes, pH7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 μM DTT andwith protease inhibitor cocktail). After 15 min of incubation,10 μL of Nonidet-P40 (10%) was added, and the mixture wasvortexed for 15 s. Thereafter, the lysate was repeatedly centri-fuged for 10 min (1000g, 4 °C) to separate the nuclear from thecytosolic fraction. The pellet was resuspended in 100 μL ofbuffer B (20 mM Hepes, pH 7.9, 0.4 M NaCl, 1 mM EDTA,1 mM EGTA, 1 μM DTT, and with protease inhibitor cocktail)and incubated on ice for 15 min, and the mixture was vortexedfor 10–15 s every 2 min. After a final centrifugation step for10 min (20 000g, 4 °C), the supernatant was collected as thenuclear extract. Protein expression was determined by Westernblots, and the band intensity from the Western blots was ana-lyzed and quantified by Image pro plus (IPP) software.
Knockdown of Nrf2 expression by shRNA
The shRNA (shNrf2-842) targeting rat Nrf2 gene and shRNAwith scrambled sequence (shNT) as control were used for Nrf2knockdown experiments.33,37 Exponentially growing cells weretransfected with different shRNAs using the GeneTran IIItransfection reagent according to the manufacturer’s instruc-tion. After 48 h of transfection, the cells were maintained inDMEM containing 10% FBS, 2 mM glutamine, 100 units perml penicillin/streptomycin at 37 °C in a humidified atmos-phere of 5% CO2 and selected by supplementation with 0.5 mgml−1 of G418. Knockdown of the Nrf2 expression in the cellswas analyzed by Western blotting.
Determination of clog P and polar surface area (PSA) of HT
The clog P of HT was obtained by determining its partitioncoefficient between n-octanol and water according to the pub-lished procedure and o-cresol was used as a positive control.49
The octanol–water partition coefficient of HT was determinedas the follows. Approximately 100 mL of n-octanol and 100 mlof distilled water were thoroughly mixed in a 500 mL separa-
tory funnel to obtain mutual saturation. To an Erlenmeyerflask containing approximately 5 mg of HT in 10 ml of water-saturated octanol, the same volume of octanol-saturated waterwas added. The flask was then shaken for 3 h at 25 °C to reachequilibrium. The two phases were separated by a separatingfunnel. Aliquots of water phase (100 μL) and octanol phase(100 μL) were transferred to cuvettes containing 1900 μL ofmethanol, and the concentrations of HT in the octanol phase(C1) and water phase (C2) were quantified by measuring theabsorbance at 281 nm and comparing it to an established cali-bration curve of HT. The clog P value was calculated by theequation: clog P = log(C1/C2). The PSA value of HT was calcu-lated online via the website: http://www.molinspiration.com.
Statistics
Data are presented as mean ± S. E. Statistical differencesbetween two groups were assessed by the Students t-test. Com-parisons among multiple groups were performed using one-way analysis of variance (ANOVA), followed by a post hocScheffe test. P < 0.05 was used as the criterion for statisticalsignificance.
Conclusions
In summary, we have demonstrated that HT is effective in pre-venting oxidative stress-induced PC12 cell damage. This pro-tection may involve its capability in both directly scavengingfree radicals and activating endogenous cellular antioxidantdefense system. Our discovery brings deep insights into under-standing the molecular mechanism underlying the cytoprotec-tive action of HT and the beneficial effects of dietary intake ofolive oil. This protective mechanism could lead to the designand development of novel small molecules as potential neuro-protective agents.
Acknowledgements
The authors appreciated Prof. Kun Gao at Lanzhou Universityand her group member for the assistance of HPLC analysis.The financial supports from the Lanzhou University (theFundamental Research Funds for the Central Universities,lzujbky-2014-56) and the Natural Science Foundation of GansuProvince (145RJZA225) are greatly acknowledged. The authorsconfirm that there are no conflicts of interest.
References
1 H. Sauer, M. Wartenberg and J. Hescheler, Cell Physiol.Biochem., 2001, 11, 173–186.
2 M. Kobayashi and M. Yamamoto, Adv. Enzyme Regul., 2006,46, 113–140.
3 K. Itoh, N. Wakabayashi, Y. Katoh, T. Ishii, K. Igarashi,J. D. Engel and M. Yamamoto, Genes Dev., 1999, 13, 76–86.
4 T. Suzuki, H. Motohashi and M. Yamamoto, Trends Phar-macol. Sci., 2013, 34, 340–346.
5 K. J. Barnham, C. L. Masters and A. I. Bush, Nat. Rev. DrugDiscovery, 2004, 3, 205–214.
6 J. A. Klein and S. L. Ackerman, J. Clin. Invest., 2003, 111,785–793.
7 A. J. Wilson, J. K. Kerns, J. F. Callahan and C. J. Moody,J. Med. Chem., 2013, 56, 7463–7476.
8 S. Magesh, Y. Chen and L. Hu, Med. Res. Rev., 2012, 32,687–726.
9 G. Blekas, C. Vassilakis, C. Harizanis, M. Tsimidou andD. G. Boskou, J. Agric. Food Chem., 2002, 50, 3688–3692.
10 R. Malheiro, P. Mendes, F. Fernandes, N. Rodrigues,A. Bento and J. A. Pereira, Food Funct., 2014, 5, 3132–3142.
11 M. Brenes, A. Garcia, P. Garcia, J. J. Rios and A. Garrido,J. Agric. Food Chem., 1999, 47, 3535–3540.
12 S. Martin-Pelaez, M. I. Covas, M. Fito, A. Kusar andI. Pravst, Mol. Nutr. Food Res., 2013, 57, 760–771.
13 S. Granados-Principal, J. L. Quiles, C. L. Ramirez-Tortosa,P. Sanchez-Rovira and M. C. Ramirez-Tortosa, Nutr. Rev.,2010, 68, 191–206.
14 F. Visioli, G. Bellomo and C. Galli, Biochem. Biophys. Res.Commun., 1998, 247, 60–64.
15 A. Chimento, I. Casaburi, C. Rosano, P. Avena, A. De Luca,C. Campana, E. Martire, M. F. Santolla, M. Maggiolini,V. Pezzi and R. Sirianni, Mol. Nutr. Food Res., 2014, 58,478–489.
16 L. J. Sun, C. Luo and J. K. Liu, Food Funct., 2014, 5, 1909–1914.
17 H. Zrelli, M. Matsuoka, S. Kitazaki, M. Zarrouk andH. Miyazaki, Eur. J. Pharmacol., 2011, 660, 275–282.
18 H. Zrelli, M. Matsuoka, S. Kitazaki, M. Araki, M. Kusunoki,M. Zarrouk and H. Miyazaki, J. Agric. Food Chem., 2011, 59,4473–4482.
19 C. St-Laurent-Thibault, M. Arseneault, F. Longpre andC. Ramassamy, Curr. Alzheimer Res., 2011, 8, 543–551.
20 S. Schaffer, W. E. Muller and G. P. Eckert, Pharmacol. Res.,2010, 62, 322–327.
21 J. A. Gonzalez-Correa, M. D. Navas, J. A. Lopez-Villodres,M. Trujillo, J. L. Espartero and J. P. De La Cruz, Neurosci.Lett., 2008, 446, 143–146.
22 R. Capasso, A. Evidente, S. Avolio and F. Solla, J. Agric. FoodChem., 1999, 47, 1745–1748.
23 J. G. Fang, M. Lu, L. P. Ma, L. Yang, L. M. Wu and Z. L. Liu,Chin. J. Chem., 2002, 20, 1313–1318.
24 K. M. Boatright and G. S. Salvesen, Curr. Opin. Cell Biol.,2003, 15, 725–731.
25 Y. M. Go and D. P. Jones, Crit. Rev. Biochem. Mol. Biol.,2013, 48, 173–181.
26 A. Bindoli, J. M. Fukuto and H. J. Forman, Antioxid. RedoxSignaling, 2008, 10, 1549–1564.
27 T. Satoh, S. R. McKercher and S. A. Lipton, Free RadicalBiol. Med., 2013, 65C, 645–657.
28 L. Zhang, D. Duan, Y. Liu, C. Ge, X. Cui, J. Sun and J. Fang,J. Am. Chem. Soc., 2014, 136, 226–233.
29 H. Pajouhesh and G. R. Lenz, NeuroRx, 2005, 2, 541–553.
30 B. Halliwell and J. Gutteridge, Free Radicals in Biology andMedicine, Oxford University Press, 4th edn, 2007.
31 C. R. Freed, P. E. Greene, R. E. Breeze, W. Y. Tsai,W. DuMouchel, R. Kao, S. Dillon, H. Winfield, S. Culver,J. Q. Trojanowski, D. Eidelberg and S. Fahn, N.Engl. J. Med., 2001, 344, 710–719.
32 S. L. Albarracin, B. Stab, Z. Casas, J. J. Sutachan,I. Samudio, J. Gonzalez, L. Gonzalo, F. Capani, L. Moralesand G. E. Barreto, Nutr. Neurosci., 2012, 15, 1–9.
33 J. Yao, C. Ge, D. Duan, B. Zhang, X. Cui, S. Peng, Y. Liu andJ. Fang, J. Agric. Food Chem., 2014, 62, 5507–5518.
34 L. W. Zhang, D. Z. Duan, X. M. Cui, J. Y. Sun andJ. G. Fang, Tetrahedron, 2013, 69, 15–21.
35 W. Cai, L. Zhang, Y. Song, B. Wang, B. Zhang, X. Cui,G. Hu, Y. Liu, J. Wu and J. Fang, Free Radical Biol. Med.,2012, 52, 257–265.
36 B. Zhang, C. Ge, J. Yao, Y. Liu, H. Xie and J. Fang, J. Am.Chem. Soc., 2015, 137, 757–769.
37 J. Yao, B. Zhang, C. Ge, S. Peng and J. Fang, J. Agric. FoodChem., 2015, 63, 1521–1531.
38 M. A. Martin, S. Ramos, A. B. Granado-Serrano,I. Rodriguez-Ramiro, M. Trujillo, L. Bravo and L. Goya, Mol.Nutr. Food Res., 2010, 54, 956–966.
39 L. Zhu, Z. Liu, Z. Feng, J. Hao, W. Shen, X. Li, L. Sun,E. Sharman, Y. Wang, K. Wertz, P. Weber, X. Shi and J. Liu,J. Nutr. Biochem., 2010, 21, 1089–1098.
40 E. Gallardo, A. Madrona, R. Palma-Valdes, M. Trujillo,J. L. Espartero and M. Santiago, RSC Adv., 2014, 4, 61086–61091.
41 T. Nguyen, C. S. Yang and C. B. Pickett, Free Radical Biol.Med., 2004, 37, 433–441.
42 T. Satoh, K. Kosaka, K. Itoh, A. Kobayashi, M. Yamamoto,Y. Shimojo, C. Kitajima, J. Cui, J. Kamins, S. Okamoto,M. Izumi, T. Shirasawa and S. A. Lipton, J. Neurochem.,2008, 104, 1116–1131.
43 Y. C. Chan, A. S. M. Ali, M. Khairuddean, K. Y. Khaw,V. Murugaiyah and A. Basiri, Chin. Chem. Lett., 2013, 24,609–612.
44 Y. Liu, D. Duan, J. Yao, B. Zhang, S. Peng, H. Ma, Y. Songand J. Fang, J. Med. Chem., 2014, 57, 5203–5211.
45 D. Duan, B. Zhang, J. Yao, Y. Liu and J. Fang, Free RadicalBiol. Med., 2014, 70, 182–193.
46 D. Duan, B. Zhang, J. Yao, Y. Liu, J. Sun, C. Ge, S. Peng andJ. Fang, Free Radical Biol. Med., 2014, 69, 15–25.
47 W. Cai, B. Zhang, D. Duan, J. Wu and J. Fang, Toxicol. Appl.Pharmacol., 2012, 262, 341–348.
48 G. L. Ellman, Arch. Biochem. Biophys., 1959, 82, 70–77.49 J. Sangster, J. Phys. Chem. Ref. Data, 1989, 18, 1111–