

Biological Journal of the Linnean Society
2007
90
319ndash348 With 3 figures
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
319
Blackwell Publishing LtdOxford UKBIJBiological Journal of the Linnean Society0024-4066copy 2007 The Linnean Society of London 200790bullbull319348Original Article
POLYDOMY IN ANTSG DEBOUT
ET AL
Corresponding author E-mail gdeboutueaacuk
Polydomy in ants what we know what we think we know and what remains to be done
GABRIEL DEBOUT
1
BERTRAND SCHATZ
2
MARIANNE ELIAS
3
and DOYLE MCKEY
2
1
CEEC
minus
School of Biological Sciences University of East Anglia Norwich NR4 7TJ UK
2
Centre drsquoEcologie Fonctionnelle et Evolutive
minus
CNRS 1919 route de Mende 34293 Montpellier Cedex 5 France
3
Department of Biological and Environmental Sciences University of Helsinki PO Box 65 FIN-00014 Helsinki Finland
Received 14 March 2005 accepted for publication 1 March 2006
The correct identification of colony boundaries is an essential prerequisite for empirical studies of ant behaviour andevolution Ant colonies function at various organizational levels and these boundaries may be difficult to assessMoreover new complexity can be generated through the presence of spatially discrete subgroups within a more orless genetically homogeneous colony a situation called polydomy A colony is polydomous only if individuals (workersand brood) of its constituent nests function as a social and cooperative unit and are regularly interchanged amongnests This condition was previously called polycalic and the term polydomy was used in a broader sense for a groupof daughter nests of the same mother colony (implying limited female dispersal) without regard to whether these dif-ferent nests continued to exchange individuals We think that this distinction between lsquopolycalyrsquo and lsquopolydomyrsquo con-cerns two disparate concepts We thus prefer the narrower definition of polydomy which groups individuals thatinteract socially Does this new level of organization affect the way in which natural selection acts on social traitsHere after examining the history of terms we review all ant species that have been described as expressing poly-domous structures We show that there is no particular syndrome of traits predictably associated with polydomy Wedetail the existing theoretical predictions and empirical results on the ecology of polydomy and the impact of poly-domy on social evolution and investment strategies while carefully distinguishing monogynous from polygynousspecies Finally we propose a methodology for future studies and offer ideas about what remains to be donecopy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
ADDITIONAL KEYWORDS
ecological constraints ndash Formicidae ndash number of queens ndash social evolution
INTRODUCTION
In social bees and wasps each colony has a single nestHowever in ants one colony may occupy either onenest (ie monodomy) or several socially connected butspatially separated nests (ie polydomy) (Houmllldobler ampWilson 1977) A colony could then be defined as agroup of related workers and associated reproduc-tives and one or more nests are the structures thathouse the colony The occurrence of polydomy in ants(as in termites Roisin Pasteels amp Braekman 1986Adams amp Levings 1987 Bulmer Adams amp Traniello2001) is perhaps related to the fact that their workers
are flightless facilitating connections of colony units(eg transport of brood) in discrete nest sites by use oftrails
Unfortunately much confusion exists concerningexact meanings of the words lsquonestrsquo lsquopolydomyrsquo andlsquopolycalyrsquo and problems with the terminology providean unavoidable source of errors Earlier work centredmainly on
Formica
was followed by studies demon-strating that a large number of ant species should beconsidered as polydomous according to the initial def-inition by Forel (1874) (ie more than one nest withbrood andor queen absent in at least one nest) Recentadvances in our knowledge of ant biology make it nec-essary to establish a new clear and unifying definitionof polydomy that is more generally valid and better
320
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
differentiates polydomous and monodomous coloniesAfter reviewing historical meanings of lsquopolydomyrsquolsquopolycalyrsquo and associated descriptive terms wepresent a synthetic review of published informationdiscussing the different hypotheses that have beenproposed to explain the existence and pattern of dis-tribution of this complex social trait We examine cor-relations between polydomy and other traits in anattempt to determine whether one or more character-istic syndromes of polydomy can be recognized
I
NTEREST
OF
STUDYING
POLYDOMY
The correct identification of colony boundaries is anessential prerequisite for empirical studies of socialinsect behaviour and evolution Testing hypothesesabout themes such as kin selection sex allocation andlevels of selection requires identifying colony bound-aries For sex allocation the colony is the basic unitupon which theory is based The presence of spatiallydiscrete subgroups within a more or less geneticallyhomogeneous colony is of prime interest because itmight permit separation of effects of queenndashworkerconflict on sex allocation from those of other poten-tially confounding factors However polydomy gener-ates new complexity because the social community of acolony is dispatched over several places and in severalunits exacerbating allocation conflicts that underlierelations within a colony Does this new level of orga-nization affect the way in which natural selection actson social traits Finally another reason to examinepolydomy is the frequent ecological success of poly-domous species or societies
All these reasons reveal a strong need for clearempirical studies and theoretical predictions to under-stand the causes and consequences of polydomy inants Empirical studies should also focus on how vari-ation in other traits (eg the number of queens) mightmodify theoretical predictions Species in which poly-domy is variable within or among populations may beespecially promising model systems for exploringselective pressures acting on this and associatedtraits
D
EFINITIONS
OF
POLYDOMY
In this review we introduce a unifying terminology todescribe polydomous structures of colonies of antsalthough it should be kept in mind that the definitionincludes several biological phenomena showing moreor less continuous variation We propose here to definelsquopolydomyrsquo as an arrangement of an ant colony in atleast two spatially separated nests The spatial sepa-ration between two nests should be obviously largerthan the usual distance between two nest chambers inthe core nest structure (see below) Second we con-
sider as a nest any structure that houses workers andbrood (essentially larvae and young pupae becauseeggs are not usually carried between nests probablydue to being too vulnerable) regardless of the numberof reproductive females in the structure (zero one ormore) The presence of a queen is not a fundamentalcriterium for the perenniality of a structure becausenew workers can be recruited to a queenless nest bythe rearing of brood from first-instar larvae and pupaetransported to it The presence of brood is fundamen-tal because it induces the expression of behaviourtypical of brood care and provisioning nest site main-tenance and defence and the renewal of generationsThe more or less complex network of communicationbetween the different nests including transport ofbrood from queenright to queenless nests argues for adefinition of polydomy that does not exclude monogy-nous species as did the definition of polycaly by Forel(1874)
Most ant species are multicolonial (ie populationsconsist of entities that function largely independentlyBourke amp Franks 1995) and these include both mon-odomous and polydomous species However in somecases notably introduced species some populationsare unicolonial (ie the constituent nests of an entirepopulation interact frequently and non-aggressivelywith each other) Unicoloniality is associated withvery low genetic differentiation between nests theentire population functions as a single huge poly-domous colony (Passera 1994 Reuter
et al
2001Tsutsui amp Case 2001 Giraud Pedersen amp Keller2002 Elias Rosengren amp Sundstroumlm 2005) Unicolo-nial societies represent a clear and distinct mode ofcolony structure (Keller 1995 Tsutsui
et al
2000)but this strategy is unstable in the long term surelybeing linked to a stage of lsquoestablishmentrsquo followingintroduction of the species into a new region (Keller1995) By contrast multicolonial societies include agreat range of variation along a continuum betweenmonodomy and polydomy
Polydomy is sometimes a seasonal phenomenon(Table 1) Generally in such cases a colony overwin-ters in one nest (rarely more than one) which thenfractionates into two or more units occupying differentnest sites during the active season coalescing onceagain the following winter (Higashi 1979 Alloway
et al
1982 MacKay amp MacKay 1984 RosengrenCherix amp Pamilo 1985 Herbers amp Grieco 1994) Insome species (eg
Formica uralensis
) the ants dispersebefore winter in numerous hibernation clusters out-side the moundnest probably as a risk-reducing strat-egy adapted to the high ground water level of theswamp habitat of the species (Rosengren amp Pamilo1983) Moreover a polydomous colony can contain oneor several queens and when the colony is polygynousqueens can be present in all nest units or only in some
POLYDOMY IN ANTS
321
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Tab
le 1
Lis
t of
th
e an
t sp
ecie
s (H
ymen
opte
ra
For
mic
idae
) sh
owin
g (o
blig
ate
or f
acu
ltat
ive)
pol
ydom
ous
colo
nia
l st
ruct
ure
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
PO
NE
RIN
AE
Am
blyo
poni
niA
mbl
yopo
ne p
alli
pes
ET
AE
NR
F 16
1T
161
C16
1
F16
1n
Am
blyo
poni
niP
rion
opel
ta a
mab
ilis
NE
O (C
entr
al A
mer
ica)
109
TR
RF
109
T10
9N
109
NF
109
n10
9
Am
blyo
poni
niP
rion
opel
ta m
odes
taN
EO
(Cen
tral
Am
eric
a)10
9T
RR
F 10
9T
109
N10
9N
109
F10
9n
109
Ect
atom
min
iP
arap
oner
a cl
avat
aN
EO
41T
RR
FA
41
D41
F41
n 41
Odo
ntom
achi
niO
dont
omac
hus
may
iN
EO
(Sou
th A
mer
ica)
124
TR
RF
124
A12
4C
124
VS
124
F12
4n
124
Pon
erin
iH
ypop
oner
a bo
ndro
itiE
T)napaJ(
LAP
OA
174
T17
4N
174
VS
174
F17
4n
174
Pon
erin
iP
achy
cond
y la
ber
thou
diE
TH
(Sou
th A
fric
a)42
T
EM
ZT
42N
42V
S 13
4F
134
n13
4
Pon
erin
iP
achy
cond
y la
hot
tent
ota
ET
H (S
outh
Afr
ica)
42
TE
MZ
T42
N42
F
42n
Pon
erin
iP
achy
cond
yla
goel
dii
NE
O (
Sout
h A
mer
ica)
124
TR
RF
124
A12
4C
124
N12
4F
124
n12
4
PSE
UD
OM
YR
ME
CIN
AE
Pse
udom
yrm
ecin
iP
seud
omyr
mex
eje
ctus
NE
A 10
0S
TO
AA
100
N10
0
O
100
n10
0
Pse
udom
yrm
ecin
iP
seud
omyr
mex
pal
lidus
NE
A 10
0S
TO
AA
100
N10
0
O
100
n10
0
Pse
udom
yrm
ecin
iP
seud
omyr
mex
sem
inol
eN
EA
100
ST
OA
A10
0N
100
O
10
0n
100
Pse
udom
yrm
ecin
iP
seud
omyr
mex
ven
efic
aN
EO
(Cen
tral
Am
eric
a)90
TR
RF
172
A90
N90
D90
O
90n
90
Pse
udom
yrm
ecin
iT
etra
pone
ra s
p P
SW-8
0 ne
ar a
ttenu
ata
TS
)aisAtsae -htuos(
IR
OR
F 14
A14
N14
F
14n
14
MY
RM
ICIN
AE
Cat
aula
cini
Cat
aula
cus
catu
volc
us
FN
NA
FR
RT
IR
OC
atau
laci
niC
atau
lacu
s gu
inee
nsis
RT
)ac irfAla rtne
Camptse
W(H
TE
RF
1T
1N
1N
49F
1
n
1
Cat
aula
cini
Cat
aula
cus
mck
eyi
RT
)acirfAlartne
C(H
TE
RF
117
A11
7N
117
N11
7F
44n
44
Cat
aula
cini
Cat
aula
cus
mut
icus
F
NN
AF
RR
TI
RO
Cep
halo
tini
Cep
halo
tes
atra
tus
NE
O 28
TR
RF
A28
N28
D28
F28
n28
Cep
halo
tini
Cep
halo
tes
min
utus
NE
O 10
9T
RR
FA
109
N10
9
F10
9n
109
Cep
halo
tini
Cep
halo
tes
umbr
acul
atus
NE
O 10
9T
RR
FA
109
E10
9
F10
9n
109
Cre
mat
ogas
trin
iC
rem
atog
aste
r af
rica
naE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
112
F11
2n
112
Cre
mat
ogas
trin
iC
rem
atog
aste
r br
evis
pino
saN
EO
(So
uth
Am
eric
a)41
TR
RF
A41
C41
D41
F41
n41
Cre
mat
ogas
trin
iC
rem
atog
aste
r cl
ariv
entr
isE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
112
F49
112
n11
2
Cre
mat
ogas
trin
iC
rem
atog
aste
r de
pres
saE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
49 1
12F
49 1
12n
112
Cre
mat
ogas
trin
iC
rem
atog
aste
r ga
bone
nsis
ET
H (
Cen
tral
Afr
ica)
50T
RR
FA
50C
50D
50F
50n
50
Cre
mat
ogas
trin
iC
rem
atog
aste
r he
lioph
ilaE
TH
(W
est A
fric
a)53
TR
OA
53A
53N
53D
53F
53n
53
Cre
mat
ogas
trin
iC
rem
atog
aste
r im
pres
saE
TH
(W
est A
fric
a)53
TR
OA
53A
53N
53D
53F
53n
53
Cre
mat
ogas
trin
iC
rem
atog
aste
r la
evis
NE
O (
Sout
h A
mer
ica)
167
TR
RF
A16
7N
167
D16
7
n16
7
Cre
mat
ogas
trin
iC
rem
atog
aste
r lim
ata
para
biot
ica
NE
O (
Sout
h A
mer
ica)
124
TR
RF
124
A41
C41
D41
F41
n41
Cre
mat
ogas
trin
iC
rem
atog
aste
r lo
ngis
pina
F
RR
T)acire
mAlartne
C(O
EN
F7
n7
Cre
mat
ogas
trin
iC
rem
atog
aste
r sc
utel
lari
sPA
L +
OR
I 117
TE
MZ
M11
7E
117
VS
117
F11
7n
117
Cre
mat
ogas
trin
iC
rem
atog
aste
r st
riat
ula
ET
H (
Cen
tral
Afr
ica)
112
TR
RF
A11
2C
112
D49
112
F11
2n
112
Lep
toth
orac
ini
Car
dioc
ondy
la e
mer
yiE
TH
+ A
ntill
a 13
0P
AN
AH
T12
9N
129
U12
9O
(uni
col)
129
n12
9
Lep
toth
orac
ini
Car
dioc
ondy
la n
uda
AA
S +
Mad
agas
car 36
130
PA
NA
H 36
T12
9N
129
U12
9O
(uni
col)
129
n12
9
Lep
toth
orac
ini
Car
dioc
ondy
la w
roug
hton
iiH
OL
+ In
dia
130
PA
NA
HT
129
N12
9U
129
O(u
nico
l)12
9n
129
Lep
toth
orac
ini
Lep
toth
orax
am
bigu
usE
TA
EN
TF
77A
142
N77
S5
F(s
ize-
dpdt
)(s
ize-
dpdt
)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)5
77
Lep
toth
orac
ini
Lep
toth
orax
cur
visp
inos
usE
TA
EN
TF
5 7
7A
154
N15
4S
5F
5 1
54 1
55y y y y y yy
Lep
toth
orac
ini
Lep
toth
orax
long
ispi
nosu
sE
TA
EN
TF
5 7
1M
78N
5 7
1S
5F
55
71
72
78
Lep
toth
orac
ini
Lep
toth
orax
nyl
ande
riE
T) yn a
mreG(
LAP
TF
68T
68N
68N
68F
6868
Lep
toth
orac
ini
Lep
toth
orax
pill
agen
sE
T)
AS
Un re tsae -htr o
N (A
EN
TF
75T
73
S73
F73
73
Lep
toth
orac
ini
Lep
toth
orax
tube
roin
terr
rupt
usPA
L (
Eur
ope)
128
TE
OA
128
T12
8C
N12
8F
9912
8
Lep
toth
orac
ini
Pro
tom
ogna
thus
a
mer
ican
usE
T)
AS
Unr etsae-ht ro
N( ( ( (
AE
NT
F54
60
T54
60
NS
54 6
0F
54n
54 6
0
a
b b
c
Myr
mic
arii
niM
yrm
icar
ia e
umen
oide
sE
TH
W
est amp
Cen
tral
Afr
ica)
99 1
05T
RO
AT
99C
99D
99F
99n
99
Myr
mic
arii
niM
yrm
icar
ia o
paci
vent
ris
ET
H
Cen
tral
amp S
outh
Afr
ica)
99T
RO
AT
99C
(gal
leri
es)
99D
99F
99n
99
Myr
mic
ini
Myr
mic
a pu
ncti
vent
ris
ET
AE
NT
F 6
T6
N15
3 4
5 6
N6
F45
153
45
73
6
Myr
mic
ini
Myr
mic
a ru
gino
dis
(mic
rogy
na f
orm
)PA
L
Eur
ope)
168
TE
TF
168
T16
8
N16
8F
168
M
yrm
icin
iM
yrm
ica
sulc
inod
isA
OE
T)eporu
E(L
APT
132
C13
2N
Fn
132
Och
etom
yrm
ecin
iW
asm
anni
a au
ropu
ncta
taH
OL
+ C
amer
oon
(int
rodu
ced
rang
e)13
0PA
NA
HT
129
N12
9U
129
O(u
nico
l)12
9n
129
Associa
ted
gyny
Colony
size
P13
3 o
r M
161
1
161
133
87
109
133
3
85 1
09 1
33
124
4
124
100
2
100
100
2
100
100
2
100
90 9
5
8 90
84
14
5 14
1
6 1
44
3 44
28
5 28
109
2
109
109
112
41 112
51
53
6 53
41
M10
9
M
41 1
33
4 94
133
87
M12
4
4 12
4M
174
2
174
P42
134
3
94W
42
1 42
W M P P M P M M1
P NL
NL
4
M M 3
3
3
3
8
53
M M
167
2
167
M P
112
3
112
129
129
2
36
129
77
1 15
6 1
29
154
155
286
94
154
155
77 7
8
2 73
78
94
97 9
5
128
2
128
54 6
0
2 54
99
6 94
105
99
7 99
NL
NL
45
2 94
95
4 94
132
2
94
97 1
29
4 94
97 1
17
M P P P P N
LP
M
NL
M P P P P P
322
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Phe
idol
ini
Aph
aeno
gast
er c
ocke
rell
iN
EA
(So
uthw
este
rn U
SA)14
6ST
MZ
T14
6E
146
D14
6F
146
n14
6
Phe
idol
ini
Mes
sor
barb
arus
PAL
(M
edit
erre
an z
one)
2 1
9E
RM
ZT
2 1
9C
(gal
leri
es)
2 1
9D
2F
(siz
e-dp
dt)
F (s
ize-
dpdt
)
2
Phe
idol
ini
Mes
sor
cape
nsis
ET
H (
Sout
h A
fric
a) 4
3T
EO
AT
43E
43D
43F
43n
43
Phe
idol
ini
Mes
sor
was
sman
ni
TA
OR
E)eporu
Enaenarretide
M(L
APC
66
F66
P
heid
olin
iP
heid
ole
anas
tasi
iN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
D10
9F
109
n10
9
Phe
idol
ini
Phe
idol
e ar
ieta
nsN
EO
(C
entr
al A
mer
ica)
109
TR
RF
M10
9N
109
F
109
n10
9
Phe
idol
ini
Phe
idol
e de
sert
orum
nF
DE
TZ
MR
E)
ASU
htuos(A
EN
Phe
idol
ini
Phe
idol
e m
egac
epha
laE
TH
NE
O A
US
79 1
30PA
NA
H 13
0T
129
79
N12
9U
79 1
29O
(un
icol
)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
79 1
29n
79 1
29
Phe
idol
ini
Phe
idol
e pa
llid
ula
PAL
(M
edit
erra
nean
zon
e) 55
ER
MZ
55T
55N
55N
55F
55n y
55
Phe
idol
ini
Pri
stom
yrm
ex p
unge
nsPA
L (
Japa
n)16
4T
ET
F 16
4T
164
N16
4N
164
F16
416
4
Phe
idol
ini (
)P
rocr
ypto
ceru
s la
eviv
entr
isN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
F
10
9n
109
Phe
idol
ini (
)P
rocr
ypto
ceru
s m
ayri
NE
O (
Cen
tral
amp S
outh
Am
eric
a)10
9T
RR
FA
109
N10
9
F
109
n10
9
Sol
enop
sidi
niM
onom
oriu
m d
estr
ucto
rH
AN
AP) ]egnar
evitan [aidnI (
IR
OT
129
N12
9U
129
129
n12
9
Sol
enop
sidi
niM
onom
oriu
m f
lori
cola
HA
NAP
)]egnarevitan [
aid nI(I
RO
T12
9N
129
U12
912
9n
129
Sol
enop
sidi
niM
onom
oriu
m m
inim
umN
EA
(N
orth
US
A) 16
2T
EO
AT
162
E16
2N
162
F16
2n
162
Sol
enop
sidi
niM
onom
oriu
m p
hara
onis
WW
129
130
PAN
AH
T12
9N
129
U12
9O
n12
9
Sol
enop
sidi
niSo
leno
psis
gem
inat
aN
EW
(C
entr
al A
mer
ica
[nat
ive
rang
e])
136
ER
OA
T9
136
E13
6D
136
F13
6n
136
Sol
enop
sidi
niSo
leno
psis
invi
cta
NE
A(i
ntro
duce
d ra
nge)
86
TE
OA
T9
D
F
Tet
ram
orii
niT
etra
mor
ium
acu
leat
um e
FR
RT
HT
EA
50 1
12C
50 1
12D
49 5
0 1
12F
49n
50
Tet
ram
orii
niT
etra
mor
ium
afr
ican
umn
FD
CA
FR
RT
HT
ET
etra
mor
iini
Tet
ram
oriu
m c
aesp
itum
PAL
(Eur
ope)
148
86
TE
TF
148
T14
8
N14
8F
148
n14
8
AN
EU
RE
TIN
AE
Ane
uret
us s
imon
iTS
)yle visu lc xeakna
LirS(
IR
OR
F 91
T91
N91
N91
F91
D
OL
ICH
OD
ER
INA
ED
olic
hode
rini
Dol
icho
deru
s qu
adri
punc
tatu
sPA
L (
Fran
ce)15
9T
ET
FA
159
N15
9N
O15
9 1
60n
Dol
icho
deri
niD
olic
hode
rus
f b
iden
sN
EO
(Bra
zil)
52 6
8T
RR
FA
52 1
03C
103
D
103
F10
3n
103
Dol
icho
deri
niL
iom
etop
um a
picu
latu
mn
FD
CT
ZM
RT
)A
SUtse
W(A
EN
Tap
inom
ini
Azt
eca
alfa
riN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
char
tifex
spi
riti
NE
O 52
TR
RF
A52
CD
Fn
Tap
inom
ini
Azt
eca
coer
ulei
penn
isN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
para
ensi
s bo
ndar
iN
EO
52T
RR
FA
52C
DF
nT
apin
omin
iA
ztec
a cf
lan
ugin
osa
NE
O (
Bra
zil)
121
TR
RF
A12
1C
121
D12
1F
121
n12
1
Tap
inom
ini
Azt
eca
ovat
icep
sN
EO
(B
razi
l)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
cf t
raili
NE
O (
Peru
)40
TR
RF
A40
C
40N
40F
40n
40
Tap
inom
ini
Azt
eca
trig
ona
NE
O (
Pana
ma)
3T
RR
F 3
A3
C3
D3
O
3n
3
Tap
inom
ini
Dor
ymy r
mex
g h in
sana
nF
DE
TA
OE
TA
EN
Tap
inom
ini
Irid
omyr
mex
n
itid
icep
sA
US
(Aus
tral
ia)
32E
RO
AT
32E
32D
32F
32n
32
Tap
inom
ini
Irid
omyr
mex
pur
pure
usA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
san
guin
eus
AU
S (A
ustr
alia
)11
5E
RO
A41
T11
5C
115
D41
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
vir
idia
eneu
sA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n y
115
Tap
inom
ini
Lin
epith
ema
hum
ileW
W (
intr
oduc
ed r
ange
) 87
130
PAN
AH
T12
9N
129
U12
912
987
Tap
inom
ini
Tap
inom
a m
elan
ocep
halu
mW
W 15
86
PAN
AH
130
T15
129
NU
15 1
2915
129
n15
129
Tap
inom
ini
Tap
inom
a m
inut
umR
E)ailartsu
A(S
UA
TF
76T
76N
76
F76
T
apin
omin
iT
echn
omyr
mex
alb
ipes
PA
L
(Jap
an) 1
65 1
66T
ET
FA
165
N16
5N
16
6F
166
n16
5
d
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
FO
RM
ICIN
AE
Cam
pono
tini
Cam
pono
tus
abdo
min
alis
flo
rida
nus
NE
A (
Flor
ida)
101
STM
ZT
101
E10
1D
101
F10
1n
101
Cam
pono
tini
Cam
opno
tus
brut
usE
TH
(C
entr
al A
fric
a)12
0T
RR
FT
120
N12
0N
120
49
F12
0n
120
Cam
pono
tini
Cam
pono
tus
cing
ulat
usN
EO
52T
RR
FA
52N
52
Fn
Cam
pono
tini
Cam
pono
tus
detr
itus
ET
H (
Sout
h-W
est A
fric
a)31
ER
MZ
T31
C31
D31
F31
n
31
Cam
pono
tini
Cam
pono
tus
giga
sR
T) oenro
B(I
RO
RF
137
139
T13
7 1
38 1
39C
137
138
139
D13
913
9n
137
138
139
Cam
pono
tini
Cam
pono
tus
fem
orat
usN
EO
40 1
24T
RR
F 12
4A
41C
124
D41
F41
n41
19
66
79 1
29
55
3
83
164
7
164
109
1
109
109
129
129
97
8 9
7
136
9 5
9 9
5 9
7
7 86
112
95
6 94
86
M
4
P
P M W M M1
P P
P P P M M
M NL
91
2 91
159
3
159
103
52
52
121
40
3 40
3
5 3
32
11
5
7 94
86
115
115
87 9
7 1
29
8
P 15
129
3
15
76
3 76
165
5
166
101
120
52
31
6
31
M
137
138
139
5
139
40 4
1
M NL
M
M M
7
M P P3
P P P P P NL
P 5
5
M P P
Associa
ted
gyny
Colony
size
Cam
pono
tini
Cam
pono
tus
herc
ulan
eus
HO
L 16
8 8
6E
RT
F 16
8T
168
D
168
F16
8n
168
Cam
pono
tini
Cam
pono
tus
impr
essu
sN
EA
(Fl
orid
a)17
1T
ET
F 17
1A
171
N17
1D
171
F17
1n y
171
Cam
pono
tini
Cam
pono
tus
kius
iuen
sis
PAL
(so
uthe
rn J
apan
)89
STT
FA
89N
89
F(s
ize-
dpdt
)89
89
Cam
pono
tini
Cam
pono
tus
ligni
perd
usPA
L (
Eur
ope)
62T
ET
FT
62E
62D
62F
62n
62
Cam
pono
tini
Cam
pono
tus
mod
ocN
EA
(C
alif
orni
a)39
STT
FT
39E
39D
39F
39n
Cam
pono
tini
Cam
pono
tus
penn
sylv
anic
usn
FD
ET
AO
ET
AE
N
Cam
pono
tini
Cam
pono
tus
plan
atus
NE
A (
Flor
ida
[int
rodu
ced
ranp
e])
16ST
RF
A16
N
16
F16
n16
Cam
pono
tini
Cam
pono
tus
isp
1O
RI
(Sou
th-E
ast
Asi
a [M
alay
Arc
hipe
lago
])T
RR
FA
N57
NF
57n
57
Cam
pono
tini
Col
obop
sis
nipp
onic
usPA
L (
Japa
n)67
TE
TF
A67
N67
NF
67n
Gig
anti
opin
iG
igan
tiop
s de
stru
ctor
NE
O (
Fren
ch G
uian
a)21
TR
RF
T21
N21
NF
n21
97 171
89
62 16
57 67 21
M M M P M P M M M
6
86 1
68
2
171
2
89
4
86 9
4
94
3
16
5
57
2
21
4
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
323
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
y yFo
rmic
ini
Cat
agly
phis
alb
ican
sPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is b
icol
orPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is ib
eric
aPA
L (
Iber
ian
peni
nsul
a) 18
ER
MZ
T37
N37
D37
F37
n37
Form
icin
iF
orm
ica
aqui
loni
a (F
orm
ica
s s
tr)
PAL
(E
urop
e)(E
urop
e)
96T
ET
FT
96C
96D
29 9
6F
96n
29
Form
icin
iF
orm
ica
brun
iPA
L
24T
ET
FT
24C
(mou
nd)24
D24
F24
n24
Form
icin
iF
orm
ica
cine
rea
(Se
rvif
orm
ica
)PA
L (
Nor
th E
urop
e)34
176
TE
TF
176
T34
C
TC
(mou
nd)34
D34
F34
n34
Form
icin
iF
orm
ica
cuni
cula
ria
PAL
(Po
land
)35T
ET
FD
34F
nFo
rmic
ini
For
mic
a ex
sect
a (
Cop
tofo
rmic
a)
PAL
(N
orth
Eur
ope)
96T
ET
F 14
8 1
25 3
3T
96C
96D
96F
96n
96 9
7
Form
icin
iF
orm
ica
exse
ctoi
des
NE
A 14
4 8
6T
ET
FT
12C
12D
12F
12n
12
Form
icin
iF
orm
ica
haem
orrh
oida
lis
nF
DC
TF
TE
TA
EN
Form
icin
iF
orm
ica
imita
nsPA
L (
Rus
sia)
176
TE
TF
176
T17
6C
176
D17
6F
176
n17
6
Form
icin
iF
orm
ica
lugu
bris
(F
orm
ica
s s
tr)
PAL
(E
urop
e)96
TE
TF
T96
C96
D96
F22
23
96
n22
23
96
Form
icin
iF
orm
ica
nigr
ican
s (
For
mic
a s
str
)PA
L (
Cen
tral
Eur
ope)
25T
ET
FT
25C
25D
25F
25n
25
Form
icin
iF
orm
ica
obsc
urip
esN
EA
(C
entr
al U
SA)
26 2
7 1
16T
ET
FT
116
E11
6D
116
F11
6n
116
Form
icin
iF
orm
ica
opac
iven
tris
FT
ET
)AS
U(A
EN
T12
3
D12
3F
123
Fo
rmic
ini
For
mic
a pa
llide
fulv
a ni
tidiv
entr
isN
EA
(ea
ster
n U
SA
)152
TE
TF
T15
2C
152
F
152
n15
2
Form
icin
iF
orm
ica
para
lugu
bris
(F
orm
ica
s s
tr)
PAL
(Sw
itze
rlan
d)11
1T
ET
FT
111
C11
1D
111
n y
111
Form
icin
iF
orm
ica
perp
ilosa
NE
A 63
170
STT
FT
63 1
70C
63D
169
F63
169
Form
icin
iF
orm
ica
podz
olic
aE
TA
EN
OA
149
TC
N14
9F
nFo
rmic
ini
For
mic
a po
lyct
ena
(F
orm
ica
s s
tr)
PAL
(E
urop
e)8
22
96
TE
TF
8 9
6 1
48T
8 9
6 1
41C
8 9
6 1
41D
22 9
6 1
41F
8 9
6 1
41n
8 1
41
Form
icin
iF
orm
ica
prat
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
25 9
6T
ET
FT
25 9
6C
25 9
6D
25 9
6 1
40F
25 9
6 1
40n
25
Form
icin
iF
orm
ica
pres
sila
bris
(C
opto
form
ica
)PA
L (
Nor
th E
urop
e)12
6 9
6T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
rufa
(F
orm
ica
s s
tr)
PAL
96T
ET
F 14
8T
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
sang
uine
a (R
aptif
orm
ica
)PA
L (
Finl
and)
96T
ET
FT
96C
96S
35 9
6F
96n
96
Form
icin
iF
orm
ica
tran
skau
kasi
ca (
Serv
ifor
mic
a)
PAL
(Fi
nlan
d)96
TE
TF
T96
C96
D96
F96
n y
96
Form
icin
iF
orm
ica
trun
coru
m
(For
mic
a s
str
)PA
L (
Eur
ope)
157
96
TE
TF
148
T96
143
158
C96
157
D96
F96
157
96 1
57
Form
icin
iF
orm
ica
ulke
i (F
orm
ica
s s
tr)
FT
ET
AE
NT
123
D
123
F12
3
Form
icin
iF
orm
ica
ural
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
96T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
yess
ensi
s (
For
mic
a s
str
)PA
L (
Japa
n C
orea
)96T
ET
FT
96C
96D
96F
96n
96
Form
icin
i
Pro
form
ica
long
iset
aPA
L (
Spai
n)58
ER
MZ
T58
C58
D58
F58
n58
Form
icin
iC
ampo
noti
niP
olyr
hach
is a
rach
neR
RT
)aisen odnI(I
RO
FA
107
N10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
ellic
osa
FR
RT
)aisenodnI(I
RO
T10
7C
107
D10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
icol
orF
RR
T)aisenodnI(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is d
ives
PAL
(Ja
pan)
+ O
RI
Mal
aysi
a86
TR
RF
A37
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
achi
s do
ddi
AU
S (
Aus
tral
ia)92
TR
RF
92A
92C
92V
S 92
F92
Form
icin
iC
ampo
noti
niP
olyr
hach
is f
urca
taF
RR
T)aisenodnI(
IR
OA
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is il
laud
ata
FR
RT
)aknaLirS (
IR
OT
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is la
bori
osa
ET
H (
Cam
eroo
n)11
9 1
20T
RR
FT
119
120
C11
9 1
20N
49 1
19 1
20F
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
oest
aPA
L (
Japa
n)14
7T
ET
FA
147
N14
7N
147
F14
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
uell
eri
FR
RT
)ai sya laM(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is n
igro
pilo
saF
RR
T)ai sen odnI (
IR
OA
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is p
roxi
ma
FR
RT
)ai sen odnI(I
RO
T10
7C
107
N10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is s
chel
leri
chae
FR
RT
)aisyalaM(
IR
OA
107
N10
7N
F10
7
Las
iini
Las
ius
alie
nus
PAL
(E
urop
e)74
72
73
86
TE
TF
T73
N73
DF
73
Las
iini
Las
ius
flav
usPA
L (
Eng
land
)172
TE
TF
172
T17
2C
172
D17
2F
172
n10
7
n10
7
n10
7
n10
7
n92
n10
7
n10
7
n11
9
n14
7
n10
7
n10
7
n10
7
n y
107
73
n17
2
150
150
37
M M M P29
96
24 96 1
25
12 176
22 2
3 9
5 9
6
25 95 111 149
8 9
5 9
6 9
7 1
41
25 9
5 9
6
96 95 9
6
96 9
7
96 9
7
P P P P96
157
96 96 58 107
107
107
107
92 107
107 147
107
107
107
107
95 9
7
95 9
7
P P P P P P P M P P P P P P P P M M M M M M M M
3
150
4
150
4
37
4
24
3
7
86 9
4
7
22 9
4
5
86 9
4
6
149
8
22 1
48
8
94 1
48
5
148
8
23 9
4
4
58
4
107
6
107
3
107
7
86 9
4 1
07
5
107
2
107
118
119
2
147
1
107
4
107
4
107
5
107
5
172
4 4 4N
LM M M M M M
Las
iini
Las
ius
min
utus
nF
DC
TA
OE
TA
EN
Las
iini
Las
ius
negl
ectu
sPA
L (
Wes
t Asi
a [n
ativ
e] E
urop
e [i
ntro
duce
d]) 15
1P
AN
AH
129
151
T10
129
N12
9U
10 1
29F
10n y
129
Las
iini
Las
ius
neon
iger
NE
A (
Nor
th-E
aste
rn U
SA
) 163
TE
OA
163
162
T73
E16
2D
162
163
F73
73 1
62
Las
iini
Las
ius
saka
gam
iN
EA
(Ja
pan)
175
TE
OA
T17
5E
175
DF
175
n17
5
Las
iini
Pse
udol
asiu
s sp
1n
ON
CM
FR
RT
L
asii
niP
seud
olas
ius
sp 2
F
NC
F
RR
T
Las
iini
Pse
udol
asiu
s sp
3
FN
C
FR
RT
10 1
29
16
3
95
175
P M P P2
P2
P2
P3
324
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Oec
ophy
llini
Oec
ophy
lla lo
ngin
oda
RT
HT
ER
F 82
173
A41
82
173
C41
D41
49
O49
F 41
154
n41
Oec
ophy
llini
Oec
ophy
lla s
mar
agdi
naA
US
(A
ustr
alia
) 80 1
35E
RR
FA
41 8
0 1
35C
41 8
0D
80O
41 1
35n
41 8
0
Pla
giol
epid
ini
Ano
plol
epis
long
ipes
WW
130
PA
NA
HT
129
N12
9U
129
129
n y
129
Pla
giol
epid
ini
Pla
giol
epis
pyg
mea
PAL
(Fr
ance
) 131
TE
OA
T13
1N
131
VS
131
F13
1
Pre
nole
pidi
niP
arat
rech
ina
bour
boni
caH
AN
AP
WW
T12
9N
129
U12
912
9n
129
Pre
nole
pidi
niP
arat
rech
ina
flav
ipes
HO
L (
Eas
t Asi
a [n
ativ
e] U
SA
(int
rodu
ced)
88P
AN
AH
T88
Pre
nole
pidi
niP
arat
rech
ina
long
icor
nis
WW
130
PA
NA
HT
129
N12
9U
129
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
129
n12
9
TE
RM
ITID
AE
(Is
opte
ra)
Nas
utit
erm
itina
eN
asut
iterm
es c
orni
ger
NE
O
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es n
igri
ceps
NE
O (
Pana
ma)
(Pan
ama)
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es p
rinc
eps
AU
S (
New
Gui
nea)
142
TR
RF
A14
2C
142
F
142
n14
2
Ret
icul
iter
mit
inae
Ret
icul
iterm
es f
lavi
pes
NE
A (
USA
) 13
R
FA
13C
142
F
13n
13
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
41
7 86
94
41 8
0
12
9
6 13
0
131
2
131
129
88
12
9
4 13
0
M P P P P P14
2
M M
Associa
ted
gyny
Colony
size
a
Syn
onym
of
P a
nti
llan
a
b
syn
onym
of
Oph
talm
opon
e
c
syn
onym
of
Har
pago
xen
us
d
syn
onym
of
M
sem
iru
fus
e
syn
onym
of
Mac
rom
isch
oid
es a
cule
atu
s
f
syn
onym
of
Hyp
ocli
nea
g
syn
onym
of
Con
omyr
ma
h
now
com
bin
ed a
s
An
onyc
hom
yrm
a n
itid
icep
s
i
syn
onym
of
Col
obop
sis
A
ll r
efer
ence
s li
sted
are
in
corp
orat
ed i
n t
he
bibl
iogr
aph
y of
th
e ar
ticl
e (
1) A
ckon
or (
1981
) 98
3)
(2)
Aco
sta
Lop
ez amp
Ser
ran
o (1
995)
(3
) A
dam
s (1
990)
99
4)
(4)
Ada
ms
amp L
evin
gs (1
987)
(5)
All
oway
et a
l
(19
82)
(6) B
ansc
hba
ch
et a
l
(19
97)
(7) B
enzi
ng
(199
1) (
8) B
eye
et a
l
(19
97)
(9) B
hat
kar
amp V
inso
n (1
987)
(10
) Boo
msm
a
et a
l
(19
90)
(11)
Bra
un
Pee
ters
amp H
oumllld
oble
r (1
994)
(12
) B
rist
ow
et a
l
(19
92)
(13)
Bu
lmer
et a
l
(20
01)
(14)
Bu
sch
inge
r
et a
l
(19
94)
(15)
Bu
stos
amp C
her
ix (
1998
)(1
6) C
arli
n R
eeve
amp C
over
(19
93)
(17)
Cer
da amp
Ret
ana
(199
2) (
18)
Cer
da
et a
l
(19
94)
(19)
Cer
dan
amp P
rovo
st (
1990
) (2
0) C
eust
ers
(197
9) (
21)
Ch
agn
e B
eugn
onamp
Dej
ean
(20
00)
(22)
Ch
erix
(19
86)
(23)
Ch
erix
(19
87)
(24)
Ch
erix
amp M
adda
len
a-F
elle
r (1
987)
(25
) C
olli
ngw
ood
(198
7) (
26)
Con
way
(19
96)
(27)
Con
way
(19
97)
(28)
Cor
n (
1980
) (2
9) C
osen
s amp
Tou
ssai
nt
(198
5)
(30)
Cro
zier
P
amil
o amp
Cro
zier
(19
84)
(31)
Cu
rtis
(19
85)
(32)
Cu
shm
an
Ras
hbr
ook
amp B
eatt
ie (
1994
) (3
3)C
zech
owsk
i (1
990)
(3
4) C
zech
owsk
i (1
999)
(3
5) C
zech
owsk
i amp
Rot
kiew
icz
(199
4)
(36)
Cze
chow
ski
amp Y
amau
chi
(199
7)
(37)
Dah
bi (
1997
) (3
8) D
ahbi
amp L
enoi
r(1
998a
) (3
9) D
avid
amp W
ood
(198
0) (
40)
Dav
idso
n (
1988
) (4
1) D
avid
son
(19
97)
(42)
Dea
n (
1989
) (4
3) D
ean
amp Y
eato
n (
1993
) (4
4) D
ebou
t
et a
l
(20
03)
(45)
DeH
eer
Bac
kus
amp H
erbe
rs (
2001
) (4
6) D
ejea
n amp
Feacuten
eacuteron
(19
93)
(47)
Dej
ean
amp L
ach
aud
(199
4)
(48)
Dej
ean
et a
l
(1
993)
(4
9) D
ejea
n
et a
l
(1
994)
(5
0) D
ejea
n
Dji
eto-
Lor
don
amp D
ura
nd
(199
7) (
51)
Dej
ean
et a
l
(20
00)
(52)
Del
abie
Ben
ton
amp d
e M
edei
ros
(199
1) (
53)
Del
age-
Dar
chen
(19
74)
(54)
Del
Rio
Pes
ado
amp A
llow
ay (
1983
)(5
5) D
etra
in (
1990
) (5
6) E
lmes
(19
87)
(57)
Fed
erle
M
asch
wit
z amp
Fia
la (
1998
) (5
8) F
ern
ande
z-E
scu
dero
et a
l
(2
001)
(5
9) F
letc
her
et a
l
(1
980)
(6
0) F
oitz
ik amp
Her
bers
(20
01)
(61)
Fra
nco
eur
amp P
eacutepin
(19
78)
(62)
Gad
au
et a
l
(19
98)
(63)
Ger
st (
2001
) (6
4) G
iber
nau
amp D
ejea
n (
2001
) (6
5) G
reen
slad
e amp
Hal
lida
y (1
983)
(66
)H
arkn
ess
amp I
sham
(19
88)
(67)
Has
egaw
a (1
992)
(6
8) H
ein
ze
et a
l
(1
996)
(6
9) H
elm
s (1
999)
(7
0) H
elm
s
et a
l
(2
000)
(7
1) H
erbe
rs (
1986
) (7
2) H
erbe
rs (
1987
)(7
3) H
erbe
rs (
1989
) (7
4) p
ers
obs
ev
cite
d in
Her
bers
(19
89)
(75)
un
publ
da
ta c
ited
in
Her
bers
(19
89)
(76)
Her
bers
(19
91)
(77)
Her
bers
amp G
riec
o (1
994)
(7
8)H
erbe
rs amp
Tu
cker
(19
86)
(79)
Hof
fman
n (
1998
) (8
0) H
oumllld
oble
r (1
983)
(81
) H
oumllld
oble
r (1
984)
(82
) H
oumllld
oble
r amp
Lu
msd
en (
1980
) (8
3) H
oumllld
oble
r amp
Moumlg
lich
(19
80)
(84)
Houmll
ldob
ler
amp W
ilso
n (
1977
) (8
5) H
oumllld
oble
r amp
Wil
son
(19
86)
(86)
Houmll
ldob
ler
amp W
ilso
n (
1990
) (8
7) H
olw
ay amp
Cas
e (2
000)
(88
) Ic
hin
ose
(198
7) (
89)
Ito
Hig
ash
iamp
Mae
ta (
1988
) (9
0) J
anze
n (
1973
) (9
1) J
ayas
uri
ya amp
Tra
nie
llo
(198
5) (
92)
Joh
nso
n amp
Cro
zier
(19
98)
(93)
Kan
now
ski
(195
9) (
94)
Kas
pari
amp V
argo
(19
95)
(95)
Kel
ler
(199
1)
(96)
Kel
ler
(199
3)
(97)
var
iou
s re
fere
nce
s in
Kel
ler
(199
8)
(98)
Kel
ler
amp P
asse
ra (
1990
) (9
9) K
enn
e (1
999)
(1
00)
Kle
in (
1987
) (1
01)
Klo
tz
et a
l
(1
996)
(1
02)
Le
Mas
ne
(199
4)
(103
) L
esto
n (
1978
) (1
04)
Leacutev
ieu
x amp
Dio
man
de (
1978
) (1
05)
Lev
ieu
x (1
983)
(1
06)
Lev
ings
amp T
ran
iell
o (1
981)
(1
07)
Lie
fke
et a
l
(1
998)
(10
8) L
ongi
no
(199
1) (
109)
Lon
gin
o (2
000)
(11
0) M
abel
is (
1994
) (1
11)
Mae
der
amp C
her
ix (
2001
) (1
12)
Maj
er (
1976
) (1
13)
Mas
chw
itz
amp M
oog
(200
0) (
114)
McG
lyn
n (
1999
) (1
15)
McI
ver
(199
1) (
116)
McI
ver
amp S
teen
(19
94)
(117
) M
cKey
D (
1984
) (1
18)
Mer
cier
amp D
ejea
n (
1996
) (1
19)
Mer
cier
Len
oir
amp D
ejea
n (
1994
)(1
20)
Mer
cier
et a
l
(19
96)
(121
) M
orai
s (1
994)
(12
2) N
icke
rson
et a
l
(19
75)
(123
) O
rsquoNei
l (19
88)
(124
) O
rive
l (20
00)
(125
) P
amil
o (1
991)
(12
6) P
amil
o amp
Ros
engr
en(1
983)
(12
7) P
amil
o C
rozi
er amp
Fra
ser
(198
5) (
128)
Par
trid
ge P
artr
idge
amp F
ran
ks (
1997
) (1
29)
Pas
sera
(19
93)
(130
) P
asse
ra (
1994
) (1
31)
Pas
sera
Gil
bert
amp A
ron
(200
1)
(132
) P
eder
sen
amp B
oom
sma
(199
9)
(133
) P
eete
rs (
1993
) (1
34)
Pee
ters
amp C
rew
e (1
986)
(1
35)
Pen
g C
hri
stia
n amp
Gib
b (1
998)
(1
36)
Per
fect
o (1
994)
(1
37)
Pfe
iffe
r amp
Lin
sen
mai
r (1
998)
(13
8) P
feif
fer
amp L
inse
nm
air
(200
0) (
139)
Pfe
iffe
r amp
Lin
sen
mai
r (2
001)
(14
0) P
irk
et a
l
(20
01)
(141
) P
isar
ski
amp C
zech
owsk
i (1
990)
(1
42)
Roi
sin
et a
l
(19
86)
(143
) R
osen
gren
et a
l
(19
85)
(144
) R
owe
amp B
rist
ow (
1999
) (1
45)
Ruuml
ppel
amp H
ein
ze (
1999
) (1
46)
San
ders
amp G
ordo
n (
2000
) (1
47)
Sas
aki
Sat
oh amp
Oba
ra (
1996
) (1
48)
Sav
olai
nen
amp V
epsauml
laumlin
en (
1988
) (1
49)
Sav
olai
nen
Vep
saumllauml
inen
amp D
esli
ppe
(199
6) (
150)
Sch
mid
-Hem
pel
(198
7) (
151)
Sei
fert
(20
00)
(152
) S
mit
h-G
lase
r (1
994)
(15
3) S
nyd
er amp
Her
bers
(19
91)
(154
) S
tuar
t (1
985)
(15
5) S
tuar
t (1
987)
(15
6) S
tuar
t (1
991)
(15
7) S
un
dstr
oumlm (
1989
) (1
58)
Su
nds
troumlm
(199
3a)
(159
) T
orro
ssia
n (
1960
) (1
60)
Tor
ossi
an (
1974
) (1
61)
Tra
nie
llo
(198
2) (
162)
Tra
nie
llo
(198
9) (
163)
Tra
nie
llo
amp L
evin
gs (
1986
) (1
64)
Tsu
ji (
1988
) (1
65)
Tsu
ji amp
Yam
auch
i (1
994)
(16
6) T
suji
et a
l
(19
91)
(167
) V
asco
nce
los
amp D
avid
son
(20
00)
(168
) V
epsauml
laumlin
en
et a
l
(20
00)
(169
) W
agn
er (
1997
) (1
70)
Wag
ner
(20
00)
(171
) W
alke
r amp
Sta
mps
(19
86)
(172
) W
alof
f amp
Bla
ckit
h (
1962
) (1
73)
Way
(19
54)
(174
) Ya
mau
chi
et a
l
(19
96)
(175
) Ya
mau
chi
et a
l
(20
01)
(176
) Z
akh
arov
(19
94)
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

320
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
differentiates polydomous and monodomous coloniesAfter reviewing historical meanings of lsquopolydomyrsquolsquopolycalyrsquo and associated descriptive terms wepresent a synthetic review of published informationdiscussing the different hypotheses that have beenproposed to explain the existence and pattern of dis-tribution of this complex social trait We examine cor-relations between polydomy and other traits in anattempt to determine whether one or more character-istic syndromes of polydomy can be recognized
I
NTEREST
OF
STUDYING
POLYDOMY
The correct identification of colony boundaries is anessential prerequisite for empirical studies of socialinsect behaviour and evolution Testing hypothesesabout themes such as kin selection sex allocation andlevels of selection requires identifying colony bound-aries For sex allocation the colony is the basic unitupon which theory is based The presence of spatiallydiscrete subgroups within a more or less geneticallyhomogeneous colony is of prime interest because itmight permit separation of effects of queenndashworkerconflict on sex allocation from those of other poten-tially confounding factors However polydomy gener-ates new complexity because the social community of acolony is dispatched over several places and in severalunits exacerbating allocation conflicts that underlierelations within a colony Does this new level of orga-nization affect the way in which natural selection actson social traits Finally another reason to examinepolydomy is the frequent ecological success of poly-domous species or societies
All these reasons reveal a strong need for clearempirical studies and theoretical predictions to under-stand the causes and consequences of polydomy inants Empirical studies should also focus on how vari-ation in other traits (eg the number of queens) mightmodify theoretical predictions Species in which poly-domy is variable within or among populations may beespecially promising model systems for exploringselective pressures acting on this and associatedtraits
D
EFINITIONS
OF
POLYDOMY
In this review we introduce a unifying terminology todescribe polydomous structures of colonies of antsalthough it should be kept in mind that the definitionincludes several biological phenomena showing moreor less continuous variation We propose here to definelsquopolydomyrsquo as an arrangement of an ant colony in atleast two spatially separated nests The spatial sepa-ration between two nests should be obviously largerthan the usual distance between two nest chambers inthe core nest structure (see below) Second we con-
sider as a nest any structure that houses workers andbrood (essentially larvae and young pupae becauseeggs are not usually carried between nests probablydue to being too vulnerable) regardless of the numberof reproductive females in the structure (zero one ormore) The presence of a queen is not a fundamentalcriterium for the perenniality of a structure becausenew workers can be recruited to a queenless nest bythe rearing of brood from first-instar larvae and pupaetransported to it The presence of brood is fundamen-tal because it induces the expression of behaviourtypical of brood care and provisioning nest site main-tenance and defence and the renewal of generationsThe more or less complex network of communicationbetween the different nests including transport ofbrood from queenright to queenless nests argues for adefinition of polydomy that does not exclude monogy-nous species as did the definition of polycaly by Forel(1874)
Most ant species are multicolonial (ie populationsconsist of entities that function largely independentlyBourke amp Franks 1995) and these include both mon-odomous and polydomous species However in somecases notably introduced species some populationsare unicolonial (ie the constituent nests of an entirepopulation interact frequently and non-aggressivelywith each other) Unicoloniality is associated withvery low genetic differentiation between nests theentire population functions as a single huge poly-domous colony (Passera 1994 Reuter
et al
2001Tsutsui amp Case 2001 Giraud Pedersen amp Keller2002 Elias Rosengren amp Sundstroumlm 2005) Unicolo-nial societies represent a clear and distinct mode ofcolony structure (Keller 1995 Tsutsui
et al
2000)but this strategy is unstable in the long term surelybeing linked to a stage of lsquoestablishmentrsquo followingintroduction of the species into a new region (Keller1995) By contrast multicolonial societies include agreat range of variation along a continuum betweenmonodomy and polydomy
Polydomy is sometimes a seasonal phenomenon(Table 1) Generally in such cases a colony overwin-ters in one nest (rarely more than one) which thenfractionates into two or more units occupying differentnest sites during the active season coalescing onceagain the following winter (Higashi 1979 Alloway
et al
1982 MacKay amp MacKay 1984 RosengrenCherix amp Pamilo 1985 Herbers amp Grieco 1994) Insome species (eg
Formica uralensis
) the ants dispersebefore winter in numerous hibernation clusters out-side the moundnest probably as a risk-reducing strat-egy adapted to the high ground water level of theswamp habitat of the species (Rosengren amp Pamilo1983) Moreover a polydomous colony can contain oneor several queens and when the colony is polygynousqueens can be present in all nest units or only in some
POLYDOMY IN ANTS
321
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Tab
le 1
Lis
t of
th
e an
t sp
ecie
s (H
ymen
opte
ra
For
mic
idae
) sh
owin
g (o
blig
ate
or f
acu
ltat
ive)
pol
ydom
ous
colo
nia
l st
ruct
ure
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
PO
NE
RIN
AE
Am
blyo
poni
niA
mbl
yopo
ne p
alli
pes
ET
AE
NR
F 16
1T
161
C16
1
F16
1n
Am
blyo
poni
niP
rion
opel
ta a
mab
ilis
NE
O (C
entr
al A
mer
ica)
109
TR
RF
109
T10
9N
109
NF
109
n10
9
Am
blyo
poni
niP
rion
opel
ta m
odes
taN
EO
(Cen
tral
Am
eric
a)10
9T
RR
F 10
9T
109
N10
9N
109
F10
9n
109
Ect
atom
min
iP
arap
oner
a cl
avat
aN
EO
41T
RR
FA
41
D41
F41
n 41
Odo
ntom
achi
niO
dont
omac
hus
may
iN
EO
(Sou
th A
mer
ica)
124
TR
RF
124
A12
4C
124
VS
124
F12
4n
124
Pon
erin
iH
ypop
oner
a bo
ndro
itiE
T)napaJ(
LAP
OA
174
T17
4N
174
VS
174
F17
4n
174
Pon
erin
iP
achy
cond
y la
ber
thou
diE
TH
(Sou
th A
fric
a)42
T
EM
ZT
42N
42V
S 13
4F
134
n13
4
Pon
erin
iP
achy
cond
y la
hot
tent
ota
ET
H (S
outh
Afr
ica)
42
TE
MZ
T42
N42
F
42n
Pon
erin
iP
achy
cond
yla
goel
dii
NE
O (
Sout
h A
mer
ica)
124
TR
RF
124
A12
4C
124
N12
4F
124
n12
4
PSE
UD
OM
YR
ME
CIN
AE
Pse
udom
yrm
ecin
iP
seud
omyr
mex
eje
ctus
NE
A 10
0S
TO
AA
100
N10
0
O
100
n10
0
Pse
udom
yrm
ecin
iP
seud
omyr
mex
pal
lidus
NE
A 10
0S
TO
AA
100
N10
0
O
100
n10
0
Pse
udom
yrm
ecin
iP
seud
omyr
mex
sem
inol
eN
EA
100
ST
OA
A10
0N
100
O
10
0n
100
Pse
udom
yrm
ecin
iP
seud
omyr
mex
ven
efic
aN
EO
(Cen
tral
Am
eric
a)90
TR
RF
172
A90
N90
D90
O
90n
90
Pse
udom
yrm
ecin
iT
etra
pone
ra s
p P
SW-8
0 ne
ar a
ttenu
ata
TS
)aisAtsae -htuos(
IR
OR
F 14
A14
N14
F
14n
14
MY
RM
ICIN
AE
Cat
aula
cini
Cat
aula
cus
catu
volc
us
FN
NA
FR
RT
IR
OC
atau
laci
niC
atau
lacu
s gu
inee
nsis
RT
)ac irfAla rtne
Camptse
W(H
TE
RF
1T
1N
1N
49F
1
n
1
Cat
aula
cini
Cat
aula
cus
mck
eyi
RT
)acirfAlartne
C(H
TE
RF
117
A11
7N
117
N11
7F
44n
44
Cat
aula
cini
Cat
aula
cus
mut
icus
F
NN
AF
RR
TI
RO
Cep
halo
tini
Cep
halo
tes
atra
tus
NE
O 28
TR
RF
A28
N28
D28
F28
n28
Cep
halo
tini
Cep
halo
tes
min
utus
NE
O 10
9T
RR
FA
109
N10
9
F10
9n
109
Cep
halo
tini
Cep
halo
tes
umbr
acul
atus
NE
O 10
9T
RR
FA
109
E10
9
F10
9n
109
Cre
mat
ogas
trin
iC
rem
atog
aste
r af
rica
naE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
112
F11
2n
112
Cre
mat
ogas
trin
iC
rem
atog
aste
r br
evis
pino
saN
EO
(So
uth
Am
eric
a)41
TR
RF
A41
C41
D41
F41
n41
Cre
mat
ogas
trin
iC
rem
atog
aste
r cl
ariv
entr
isE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
112
F49
112
n11
2
Cre
mat
ogas
trin
iC
rem
atog
aste
r de
pres
saE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
49 1
12F
49 1
12n
112
Cre
mat
ogas
trin
iC
rem
atog
aste
r ga
bone
nsis
ET
H (
Cen
tral
Afr
ica)
50T
RR
FA
50C
50D
50F
50n
50
Cre
mat
ogas
trin
iC
rem
atog
aste
r he
lioph
ilaE
TH
(W
est A
fric
a)53
TR
OA
53A
53N
53D
53F
53n
53
Cre
mat
ogas
trin
iC
rem
atog
aste
r im
pres
saE
TH
(W
est A
fric
a)53
TR
OA
53A
53N
53D
53F
53n
53
Cre
mat
ogas
trin
iC
rem
atog
aste
r la
evis
NE
O (
Sout
h A
mer
ica)
167
TR
RF
A16
7N
167
D16
7
n16
7
Cre
mat
ogas
trin
iC
rem
atog
aste
r lim
ata
para
biot
ica
NE
O (
Sout
h A
mer
ica)
124
TR
RF
124
A41
C41
D41
F41
n41
Cre
mat
ogas
trin
iC
rem
atog
aste
r lo
ngis
pina
F
RR
T)acire
mAlartne
C(O
EN
F7
n7
Cre
mat
ogas
trin
iC
rem
atog
aste
r sc
utel
lari
sPA
L +
OR
I 117
TE
MZ
M11
7E
117
VS
117
F11
7n
117
Cre
mat
ogas
trin
iC
rem
atog
aste
r st
riat
ula
ET
H (
Cen
tral
Afr
ica)
112
TR
RF
A11
2C
112
D49
112
F11
2n
112
Lep
toth
orac
ini
Car
dioc
ondy
la e
mer
yiE
TH
+ A
ntill
a 13
0P
AN
AH
T12
9N
129
U12
9O
(uni
col)
129
n12
9
Lep
toth
orac
ini
Car
dioc
ondy
la n
uda
AA
S +
Mad
agas
car 36
130
PA
NA
H 36
T12
9N
129
U12
9O
(uni
col)
129
n12
9
Lep
toth
orac
ini
Car
dioc
ondy
la w
roug
hton
iiH
OL
+ In
dia
130
PA
NA
HT
129
N12
9U
129
O(u
nico
l)12
9n
129
Lep
toth
orac
ini
Lep
toth
orax
am
bigu
usE
TA
EN
TF
77A
142
N77
S5
F(s
ize-
dpdt
)(s
ize-
dpdt
)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)5
77
Lep
toth
orac
ini
Lep
toth
orax
cur
visp
inos
usE
TA
EN
TF
5 7
7A
154
N15
4S
5F
5 1
54 1
55y y y y y yy
Lep
toth
orac
ini
Lep
toth
orax
long
ispi
nosu
sE
TA
EN
TF
5 7
1M
78N
5 7
1S
5F
55
71
72
78
Lep
toth
orac
ini
Lep
toth
orax
nyl
ande
riE
T) yn a
mreG(
LAP
TF
68T
68N
68N
68F
6868
Lep
toth
orac
ini
Lep
toth
orax
pill
agen
sE
T)
AS
Un re tsae -htr o
N (A
EN
TF
75T
73
S73
F73
73
Lep
toth
orac
ini
Lep
toth
orax
tube
roin
terr
rupt
usPA
L (
Eur
ope)
128
TE
OA
128
T12
8C
N12
8F
9912
8
Lep
toth
orac
ini
Pro
tom
ogna
thus
a
mer
ican
usE
T)
AS
Unr etsae-ht ro
N( ( ( (
AE
NT
F54
60
T54
60
NS
54 6
0F
54n
54 6
0
a
b b
c
Myr
mic
arii
niM
yrm
icar
ia e
umen
oide
sE
TH
W
est amp
Cen
tral
Afr
ica)
99 1
05T
RO
AT
99C
99D
99F
99n
99
Myr
mic
arii
niM
yrm
icar
ia o
paci
vent
ris
ET
H
Cen
tral
amp S
outh
Afr
ica)
99T
RO
AT
99C
(gal
leri
es)
99D
99F
99n
99
Myr
mic
ini
Myr
mic
a pu
ncti
vent
ris
ET
AE
NT
F 6
T6
N15
3 4
5 6
N6
F45
153
45
73
6
Myr
mic
ini
Myr
mic
a ru
gino
dis
(mic
rogy
na f
orm
)PA
L
Eur
ope)
168
TE
TF
168
T16
8
N16
8F
168
M
yrm
icin
iM
yrm
ica
sulc
inod
isA
OE
T)eporu
E(L
APT
132
C13
2N
Fn
132
Och
etom
yrm
ecin
iW
asm
anni
a au
ropu
ncta
taH
OL
+ C
amer
oon
(int
rodu
ced
rang
e)13
0PA
NA
HT
129
N12
9U
129
O(u
nico
l)12
9n
129
Associa
ted
gyny
Colony
size
P13
3 o
r M
161
1
161
133
87
109
133
3
85 1
09 1
33
124
4
124
100
2
100
100
2
100
100
2
100
90 9
5
8 90
84
14
5 14
1
6 1
44
3 44
28
5 28
109
2
109
109
112
41 112
51
53
6 53
41
M10
9
M
41 1
33
4 94
133
87
M12
4
4 12
4M
174
2
174
P42
134
3
94W
42
1 42
W M P P M P M M1
P NL
NL
4
M M 3
3
3
3
8
53
M M
167
2
167
M P
112
3
112
129
129
2
36
129
77
1 15
6 1
29
154
155
286
94
154
155
77 7
8
2 73
78
94
97 9
5
128
2
128
54 6
0
2 54
99
6 94
105
99
7 99
NL
NL
45
2 94
95
4 94
132
2
94
97 1
29
4 94
97 1
17
M P P P P N
LP
M
NL
M P P P P P
322
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Phe
idol
ini
Aph
aeno
gast
er c
ocke
rell
iN
EA
(So
uthw
este
rn U
SA)14
6ST
MZ
T14
6E
146
D14
6F
146
n14
6
Phe
idol
ini
Mes
sor
barb
arus
PAL
(M
edit
erre
an z
one)
2 1
9E
RM
ZT
2 1
9C
(gal
leri
es)
2 1
9D
2F
(siz
e-dp
dt)
F (s
ize-
dpdt
)
2
Phe
idol
ini
Mes
sor
cape
nsis
ET
H (
Sout
h A
fric
a) 4
3T
EO
AT
43E
43D
43F
43n
43
Phe
idol
ini
Mes
sor
was
sman
ni
TA
OR
E)eporu
Enaenarretide
M(L
APC
66
F66
P
heid
olin
iP
heid
ole
anas
tasi
iN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
D10
9F
109
n10
9
Phe
idol
ini
Phe
idol
e ar
ieta
nsN
EO
(C
entr
al A
mer
ica)
109
TR
RF
M10
9N
109
F
109
n10
9
Phe
idol
ini
Phe
idol
e de
sert
orum
nF
DE
TZ
MR
E)
ASU
htuos(A
EN
Phe
idol
ini
Phe
idol
e m
egac
epha
laE
TH
NE
O A
US
79 1
30PA
NA
H 13
0T
129
79
N12
9U
79 1
29O
(un
icol
)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
79 1
29n
79 1
29
Phe
idol
ini
Phe
idol
e pa
llid
ula
PAL
(M
edit
erra
nean
zon
e) 55
ER
MZ
55T
55N
55N
55F
55n y
55
Phe
idol
ini
Pri
stom
yrm
ex p
unge
nsPA
L (
Japa
n)16
4T
ET
F 16
4T
164
N16
4N
164
F16
416
4
Phe
idol
ini (
)P
rocr
ypto
ceru
s la
eviv
entr
isN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
F
10
9n
109
Phe
idol
ini (
)P
rocr
ypto
ceru
s m
ayri
NE
O (
Cen
tral
amp S
outh
Am
eric
a)10
9T
RR
FA
109
N10
9
F
109
n10
9
Sol
enop
sidi
niM
onom
oriu
m d
estr
ucto
rH
AN
AP) ]egnar
evitan [aidnI (
IR
OT
129
N12
9U
129
129
n12
9
Sol
enop
sidi
niM
onom
oriu
m f
lori
cola
HA
NAP
)]egnarevitan [
aid nI(I
RO
T12
9N
129
U12
912
9n
129
Sol
enop
sidi
niM
onom
oriu
m m
inim
umN
EA
(N
orth
US
A) 16
2T
EO
AT
162
E16
2N
162
F16
2n
162
Sol
enop
sidi
niM
onom
oriu
m p
hara
onis
WW
129
130
PAN
AH
T12
9N
129
U12
9O
n12
9
Sol
enop
sidi
niSo
leno
psis
gem
inat
aN
EW
(C
entr
al A
mer
ica
[nat
ive
rang
e])
136
ER
OA
T9
136
E13
6D
136
F13
6n
136
Sol
enop
sidi
niSo
leno
psis
invi
cta
NE
A(i
ntro
duce
d ra
nge)
86
TE
OA
T9
D
F
Tet
ram
orii
niT
etra
mor
ium
acu
leat
um e
FR
RT
HT
EA
50 1
12C
50 1
12D
49 5
0 1
12F
49n
50
Tet
ram
orii
niT
etra
mor
ium
afr
ican
umn
FD
CA
FR
RT
HT
ET
etra
mor
iini
Tet
ram
oriu
m c
aesp
itum
PAL
(Eur
ope)
148
86
TE
TF
148
T14
8
N14
8F
148
n14
8
AN
EU
RE
TIN
AE
Ane
uret
us s
imon
iTS
)yle visu lc xeakna
LirS(
IR
OR
F 91
T91
N91
N91
F91
D
OL
ICH
OD
ER
INA
ED
olic
hode
rini
Dol
icho
deru
s qu
adri
punc
tatu
sPA
L (
Fran
ce)15
9T
ET
FA
159
N15
9N
O15
9 1
60n
Dol
icho
deri
niD
olic
hode
rus
f b
iden
sN
EO
(Bra
zil)
52 6
8T
RR
FA
52 1
03C
103
D
103
F10
3n
103
Dol
icho
deri
niL
iom
etop
um a
picu
latu
mn
FD
CT
ZM
RT
)A
SUtse
W(A
EN
Tap
inom
ini
Azt
eca
alfa
riN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
char
tifex
spi
riti
NE
O 52
TR
RF
A52
CD
Fn
Tap
inom
ini
Azt
eca
coer
ulei
penn
isN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
para
ensi
s bo
ndar
iN
EO
52T
RR
FA
52C
DF
nT
apin
omin
iA
ztec
a cf
lan
ugin
osa
NE
O (
Bra
zil)
121
TR
RF
A12
1C
121
D12
1F
121
n12
1
Tap
inom
ini
Azt
eca
ovat
icep
sN
EO
(B
razi
l)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
cf t
raili
NE
O (
Peru
)40
TR
RF
A40
C
40N
40F
40n
40
Tap
inom
ini
Azt
eca
trig
ona
NE
O (
Pana
ma)
3T
RR
F 3
A3
C3
D3
O
3n
3
Tap
inom
ini
Dor
ymy r
mex
g h in
sana
nF
DE
TA
OE
TA
EN
Tap
inom
ini
Irid
omyr
mex
n
itid
icep
sA
US
(Aus
tral
ia)
32E
RO
AT
32E
32D
32F
32n
32
Tap
inom
ini
Irid
omyr
mex
pur
pure
usA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
san
guin
eus
AU
S (A
ustr
alia
)11
5E
RO
A41
T11
5C
115
D41
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
vir
idia
eneu
sA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n y
115
Tap
inom
ini
Lin
epith
ema
hum
ileW
W (
intr
oduc
ed r
ange
) 87
130
PAN
AH
T12
9N
129
U12
912
987
Tap
inom
ini
Tap
inom
a m
elan
ocep
halu
mW
W 15
86
PAN
AH
130
T15
129
NU
15 1
2915
129
n15
129
Tap
inom
ini
Tap
inom
a m
inut
umR
E)ailartsu
A(S
UA
TF
76T
76N
76
F76
T
apin
omin
iT
echn
omyr
mex
alb
ipes
PA
L
(Jap
an) 1
65 1
66T
ET
FA
165
N16
5N
16
6F
166
n16
5
d
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
FO
RM
ICIN
AE
Cam
pono
tini
Cam
pono
tus
abdo
min
alis
flo
rida
nus
NE
A (
Flor
ida)
101
STM
ZT
101
E10
1D
101
F10
1n
101
Cam
pono
tini
Cam
opno
tus
brut
usE
TH
(C
entr
al A
fric
a)12
0T
RR
FT
120
N12
0N
120
49
F12
0n
120
Cam
pono
tini
Cam
pono
tus
cing
ulat
usN
EO
52T
RR
FA
52N
52
Fn
Cam
pono
tini
Cam
pono
tus
detr
itus
ET
H (
Sout
h-W
est A
fric
a)31
ER
MZ
T31
C31
D31
F31
n
31
Cam
pono
tini
Cam
pono
tus
giga
sR
T) oenro
B(I
RO
RF
137
139
T13
7 1
38 1
39C
137
138
139
D13
913
9n
137
138
139
Cam
pono
tini
Cam
pono
tus
fem
orat
usN
EO
40 1
24T
RR
F 12
4A
41C
124
D41
F41
n41
19
66
79 1
29
55
3
83
164
7
164
109
1
109
109
129
129
97
8 9
7
136
9 5
9 9
5 9
7
7 86
112
95
6 94
86
M
4
P
P M W M M1
P P
P P P M M
M NL
91
2 91
159
3
159
103
52
52
121
40
3 40
3
5 3
32
11
5
7 94
86
115
115
87 9
7 1
29
8
P 15
129
3
15
76
3 76
165
5
166
101
120
52
31
6
31
M
137
138
139
5
139
40 4
1
M NL
M
M M
7
M P P3
P P P P P NL
P 5
5
M P P
Associa
ted
gyny
Colony
size
Cam
pono
tini
Cam
pono
tus
herc
ulan
eus
HO
L 16
8 8
6E
RT
F 16
8T
168
D
168
F16
8n
168
Cam
pono
tini
Cam
pono
tus
impr
essu
sN
EA
(Fl
orid
a)17
1T
ET
F 17
1A
171
N17
1D
171
F17
1n y
171
Cam
pono
tini
Cam
pono
tus
kius
iuen
sis
PAL
(so
uthe
rn J
apan
)89
STT
FA
89N
89
F(s
ize-
dpdt
)89
89
Cam
pono
tini
Cam
pono
tus
ligni
perd
usPA
L (
Eur
ope)
62T
ET
FT
62E
62D
62F
62n
62
Cam
pono
tini
Cam
pono
tus
mod
ocN
EA
(C
alif
orni
a)39
STT
FT
39E
39D
39F
39n
Cam
pono
tini
Cam
pono
tus
penn
sylv
anic
usn
FD
ET
AO
ET
AE
N
Cam
pono
tini
Cam
pono
tus
plan
atus
NE
A (
Flor
ida
[int
rodu
ced
ranp
e])
16ST
RF
A16
N
16
F16
n16
Cam
pono
tini
Cam
pono
tus
isp
1O
RI
(Sou
th-E
ast
Asi
a [M
alay
Arc
hipe
lago
])T
RR
FA
N57
NF
57n
57
Cam
pono
tini
Col
obop
sis
nipp
onic
usPA
L (
Japa
n)67
TE
TF
A67
N67
NF
67n
Gig
anti
opin
iG
igan
tiop
s de
stru
ctor
NE
O (
Fren
ch G
uian
a)21
TR
RF
T21
N21
NF
n21
97 171
89
62 16
57 67 21
M M M P M P M M M
6
86 1
68
2
171
2
89
4
86 9
4
94
3
16
5
57
2
21
4
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
323
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
y yFo
rmic
ini
Cat
agly
phis
alb
ican
sPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is b
icol
orPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is ib
eric
aPA
L (
Iber
ian
peni
nsul
a) 18
ER
MZ
T37
N37
D37
F37
n37
Form
icin
iF
orm
ica
aqui
loni
a (F
orm
ica
s s
tr)
PAL
(E
urop
e)(E
urop
e)
96T
ET
FT
96C
96D
29 9
6F
96n
29
Form
icin
iF
orm
ica
brun
iPA
L
24T
ET
FT
24C
(mou
nd)24
D24
F24
n24
Form
icin
iF
orm
ica
cine
rea
(Se
rvif
orm
ica
)PA
L (
Nor
th E
urop
e)34
176
TE
TF
176
T34
C
TC
(mou
nd)34
D34
F34
n34
Form
icin
iF
orm
ica
cuni
cula
ria
PAL
(Po
land
)35T
ET
FD
34F
nFo
rmic
ini
For
mic
a ex
sect
a (
Cop
tofo
rmic
a)
PAL
(N
orth
Eur
ope)
96T
ET
F 14
8 1
25 3
3T
96C
96D
96F
96n
96 9
7
Form
icin
iF
orm
ica
exse
ctoi
des
NE
A 14
4 8
6T
ET
FT
12C
12D
12F
12n
12
Form
icin
iF
orm
ica
haem
orrh
oida
lis
nF
DC
TF
TE
TA
EN
Form
icin
iF
orm
ica
imita
nsPA
L (
Rus
sia)
176
TE
TF
176
T17
6C
176
D17
6F
176
n17
6
Form
icin
iF
orm
ica
lugu
bris
(F
orm
ica
s s
tr)
PAL
(E
urop
e)96
TE
TF
T96
C96
D96
F22
23
96
n22
23
96
Form
icin
iF
orm
ica
nigr
ican
s (
For
mic
a s
str
)PA
L (
Cen
tral
Eur
ope)
25T
ET
FT
25C
25D
25F
25n
25
Form
icin
iF
orm
ica
obsc
urip
esN
EA
(C
entr
al U
SA)
26 2
7 1
16T
ET
FT
116
E11
6D
116
F11
6n
116
Form
icin
iF
orm
ica
opac
iven
tris
FT
ET
)AS
U(A
EN
T12
3
D12
3F
123
Fo
rmic
ini
For
mic
a pa
llide
fulv
a ni
tidiv
entr
isN
EA
(ea
ster
n U
SA
)152
TE
TF
T15
2C
152
F
152
n15
2
Form
icin
iF
orm
ica
para
lugu
bris
(F
orm
ica
s s
tr)
PAL
(Sw
itze
rlan
d)11
1T
ET
FT
111
C11
1D
111
n y
111
Form
icin
iF
orm
ica
perp
ilosa
NE
A 63
170
STT
FT
63 1
70C
63D
169
F63
169
Form
icin
iF
orm
ica
podz
olic
aE
TA
EN
OA
149
TC
N14
9F
nFo
rmic
ini
For
mic
a po
lyct
ena
(F
orm
ica
s s
tr)
PAL
(E
urop
e)8
22
96
TE
TF
8 9
6 1
48T
8 9
6 1
41C
8 9
6 1
41D
22 9
6 1
41F
8 9
6 1
41n
8 1
41
Form
icin
iF
orm
ica
prat
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
25 9
6T
ET
FT
25 9
6C
25 9
6D
25 9
6 1
40F
25 9
6 1
40n
25
Form
icin
iF
orm
ica
pres
sila
bris
(C
opto
form
ica
)PA
L (
Nor
th E
urop
e)12
6 9
6T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
rufa
(F
orm
ica
s s
tr)
PAL
96T
ET
F 14
8T
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
sang
uine
a (R
aptif
orm
ica
)PA
L (
Finl
and)
96T
ET
FT
96C
96S
35 9
6F
96n
96
Form
icin
iF
orm
ica
tran
skau
kasi
ca (
Serv
ifor
mic
a)
PAL
(Fi
nlan
d)96
TE
TF
T96
C96
D96
F96
n y
96
Form
icin
iF
orm
ica
trun
coru
m
(For
mic
a s
str
)PA
L (
Eur
ope)
157
96
TE
TF
148
T96
143
158
C96
157
D96
F96
157
96 1
57
Form
icin
iF
orm
ica
ulke
i (F
orm
ica
s s
tr)
FT
ET
AE
NT
123
D
123
F12
3
Form
icin
iF
orm
ica
ural
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
96T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
yess
ensi
s (
For
mic
a s
str
)PA
L (
Japa
n C
orea
)96T
ET
FT
96C
96D
96F
96n
96
Form
icin
i
Pro
form
ica
long
iset
aPA
L (
Spai
n)58
ER
MZ
T58
C58
D58
F58
n58
Form
icin
iC
ampo
noti
niP
olyr
hach
is a
rach
neR
RT
)aisen odnI(I
RO
FA
107
N10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
ellic
osa
FR
RT
)aisenodnI(I
RO
T10
7C
107
D10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
icol
orF
RR
T)aisenodnI(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is d
ives
PAL
(Ja
pan)
+ O
RI
Mal
aysi
a86
TR
RF
A37
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
achi
s do
ddi
AU
S (
Aus
tral
ia)92
TR
RF
92A
92C
92V
S 92
F92
Form
icin
iC
ampo
noti
niP
olyr
hach
is f
urca
taF
RR
T)aisenodnI(
IR
OA
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is il
laud
ata
FR
RT
)aknaLirS (
IR
OT
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is la
bori
osa
ET
H (
Cam
eroo
n)11
9 1
20T
RR
FT
119
120
C11
9 1
20N
49 1
19 1
20F
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
oest
aPA
L (
Japa
n)14
7T
ET
FA
147
N14
7N
147
F14
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
uell
eri
FR
RT
)ai sya laM(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is n
igro
pilo
saF
RR
T)ai sen odnI (
IR
OA
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is p
roxi
ma
FR
RT
)ai sen odnI(I
RO
T10
7C
107
N10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is s
chel
leri
chae
FR
RT
)aisyalaM(
IR
OA
107
N10
7N
F10
7
Las
iini
Las
ius
alie
nus
PAL
(E
urop
e)74
72
73
86
TE
TF
T73
N73
DF
73
Las
iini
Las
ius
flav
usPA
L (
Eng
land
)172
TE
TF
172
T17
2C
172
D17
2F
172
n10
7
n10
7
n10
7
n10
7
n92
n10
7
n10
7
n11
9
n14
7
n10
7
n10
7
n10
7
n y
107
73
n17
2
150
150
37
M M M P29
96
24 96 1
25
12 176
22 2
3 9
5 9
6
25 95 111 149
8 9
5 9
6 9
7 1
41
25 9
5 9
6
96 95 9
6
96 9
7
96 9
7
P P P P96
157
96 96 58 107
107
107
107
92 107
107 147
107
107
107
107
95 9
7
95 9
7
P P P P P P P M P P P P P P P P M M M M M M M M
3
150
4
150
4
37
4
24
3
7
86 9
4
7
22 9
4
5
86 9
4
6
149
8
22 1
48
8
94 1
48
5
148
8
23 9
4
4
58
4
107
6
107
3
107
7
86 9
4 1
07
5
107
2
107
118
119
2
147
1
107
4
107
4
107
5
107
5
172
4 4 4N
LM M M M M M
Las
iini
Las
ius
min
utus
nF
DC
TA
OE
TA
EN
Las
iini
Las
ius
negl
ectu
sPA
L (
Wes
t Asi
a [n
ativ
e] E
urop
e [i
ntro
duce
d]) 15
1P
AN
AH
129
151
T10
129
N12
9U
10 1
29F
10n y
129
Las
iini
Las
ius
neon
iger
NE
A (
Nor
th-E
aste
rn U
SA
) 163
TE
OA
163
162
T73
E16
2D
162
163
F73
73 1
62
Las
iini
Las
ius
saka
gam
iN
EA
(Ja
pan)
175
TE
OA
T17
5E
175
DF
175
n17
5
Las
iini
Pse
udol
asiu
s sp
1n
ON
CM
FR
RT
L
asii
niP
seud
olas
ius
sp 2
F
NC
F
RR
T
Las
iini
Pse
udol
asiu
s sp
3
FN
C
FR
RT
10 1
29
16
3
95
175
P M P P2
P2
P2
P3
324
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Oec
ophy
llini
Oec
ophy
lla lo
ngin
oda
RT
HT
ER
F 82
173
A41
82
173
C41
D41
49
O49
F 41
154
n41
Oec
ophy
llini
Oec
ophy
lla s
mar
agdi
naA
US
(A
ustr
alia
) 80 1
35E
RR
FA
41 8
0 1
35C
41 8
0D
80O
41 1
35n
41 8
0
Pla
giol
epid
ini
Ano
plol
epis
long
ipes
WW
130
PA
NA
HT
129
N12
9U
129
129
n y
129
Pla
giol
epid
ini
Pla
giol
epis
pyg
mea
PAL
(Fr
ance
) 131
TE
OA
T13
1N
131
VS
131
F13
1
Pre
nole
pidi
niP
arat
rech
ina
bour
boni
caH
AN
AP
WW
T12
9N
129
U12
912
9n
129
Pre
nole
pidi
niP
arat
rech
ina
flav
ipes
HO
L (
Eas
t Asi
a [n
ativ
e] U
SA
(int
rodu
ced)
88P
AN
AH
T88
Pre
nole
pidi
niP
arat
rech
ina
long
icor
nis
WW
130
PA
NA
HT
129
N12
9U
129
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
129
n12
9
TE
RM
ITID
AE
(Is
opte
ra)
Nas
utit
erm
itina
eN
asut
iterm
es c
orni
ger
NE
O
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es n
igri
ceps
NE
O (
Pana
ma)
(Pan
ama)
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es p
rinc
eps
AU
S (
New
Gui
nea)
142
TR
RF
A14
2C
142
F
142
n14
2
Ret
icul
iter
mit
inae
Ret
icul
iterm
es f
lavi
pes
NE
A (
USA
) 13
R
FA
13C
142
F
13n
13
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
41
7 86
94
41 8
0
12
9
6 13
0
131
2
131
129
88
12
9
4 13
0
M P P P P P14
2
M M
Associa
ted
gyny
Colony
size
a
Syn
onym
of
P a
nti
llan
a
b
syn
onym
of
Oph
talm
opon
e
c
syn
onym
of
Har
pago
xen
us
d
syn
onym
of
M
sem
iru
fus
e
syn
onym
of
Mac
rom
isch
oid
es a
cule
atu
s
f
syn
onym
of
Hyp
ocli
nea
g
syn
onym
of
Con
omyr
ma
h
now
com
bin
ed a
s
An
onyc
hom
yrm
a n
itid
icep
s
i
syn
onym
of
Col
obop
sis
A
ll r
efer
ence
s li
sted
are
in
corp
orat
ed i
n t
he
bibl
iogr
aph
y of
th
e ar
ticl
e (
1) A
ckon
or (
1981
) 98
3)
(2)
Aco
sta
Lop
ez amp
Ser
ran
o (1
995)
(3
) A
dam
s (1
990)
99
4)
(4)
Ada
ms
amp L
evin
gs (1
987)
(5)
All
oway
et a
l
(19
82)
(6) B
ansc
hba
ch
et a
l
(19
97)
(7) B
enzi
ng
(199
1) (
8) B
eye
et a
l
(19
97)
(9) B
hat
kar
amp V
inso
n (1
987)
(10
) Boo
msm
a
et a
l
(19
90)
(11)
Bra
un
Pee
ters
amp H
oumllld
oble
r (1
994)
(12
) B
rist
ow
et a
l
(19
92)
(13)
Bu
lmer
et a
l
(20
01)
(14)
Bu
sch
inge
r
et a
l
(19
94)
(15)
Bu
stos
amp C
her
ix (
1998
)(1
6) C
arli
n R
eeve
amp C
over
(19
93)
(17)
Cer
da amp
Ret
ana
(199
2) (
18)
Cer
da
et a
l
(19
94)
(19)
Cer
dan
amp P
rovo
st (
1990
) (2
0) C
eust
ers
(197
9) (
21)
Ch
agn
e B
eugn
onamp
Dej
ean
(20
00)
(22)
Ch
erix
(19
86)
(23)
Ch
erix
(19
87)
(24)
Ch
erix
amp M
adda
len
a-F
elle
r (1
987)
(25
) C
olli
ngw
ood
(198
7) (
26)
Con
way
(19
96)
(27)
Con
way
(19
97)
(28)
Cor
n (
1980
) (2
9) C
osen
s amp
Tou
ssai
nt
(198
5)
(30)
Cro
zier
P
amil
o amp
Cro
zier
(19
84)
(31)
Cu
rtis
(19
85)
(32)
Cu
shm
an
Ras
hbr
ook
amp B
eatt
ie (
1994
) (3
3)C
zech
owsk
i (1
990)
(3
4) C
zech
owsk
i (1
999)
(3
5) C
zech
owsk
i amp
Rot
kiew
icz
(199
4)
(36)
Cze
chow
ski
amp Y
amau
chi
(199
7)
(37)
Dah
bi (
1997
) (3
8) D
ahbi
amp L
enoi
r(1
998a
) (3
9) D
avid
amp W
ood
(198
0) (
40)
Dav
idso
n (
1988
) (4
1) D
avid
son
(19
97)
(42)
Dea
n (
1989
) (4
3) D
ean
amp Y
eato
n (
1993
) (4
4) D
ebou
t
et a
l
(20
03)
(45)
DeH
eer
Bac
kus
amp H
erbe
rs (
2001
) (4
6) D
ejea
n amp
Feacuten
eacuteron
(19
93)
(47)
Dej
ean
amp L
ach
aud
(199
4)
(48)
Dej
ean
et a
l
(1
993)
(4
9) D
ejea
n
et a
l
(1
994)
(5
0) D
ejea
n
Dji
eto-
Lor
don
amp D
ura
nd
(199
7) (
51)
Dej
ean
et a
l
(20
00)
(52)
Del
abie
Ben
ton
amp d
e M
edei
ros
(199
1) (
53)
Del
age-
Dar
chen
(19
74)
(54)
Del
Rio
Pes
ado
amp A
llow
ay (
1983
)(5
5) D
etra
in (
1990
) (5
6) E
lmes
(19
87)
(57)
Fed
erle
M
asch
wit
z amp
Fia
la (
1998
) (5
8) F
ern
ande
z-E
scu
dero
et a
l
(2
001)
(5
9) F
letc
her
et a
l
(1
980)
(6
0) F
oitz
ik amp
Her
bers
(20
01)
(61)
Fra
nco
eur
amp P
eacutepin
(19
78)
(62)
Gad
au
et a
l
(19
98)
(63)
Ger
st (
2001
) (6
4) G
iber
nau
amp D
ejea
n (
2001
) (6
5) G
reen
slad
e amp
Hal
lida
y (1
983)
(66
)H
arkn
ess
amp I
sham
(19
88)
(67)
Has
egaw
a (1
992)
(6
8) H
ein
ze
et a
l
(1
996)
(6
9) H
elm
s (1
999)
(7
0) H
elm
s
et a
l
(2
000)
(7
1) H
erbe
rs (
1986
) (7
2) H
erbe
rs (
1987
)(7
3) H
erbe
rs (
1989
) (7
4) p
ers
obs
ev
cite
d in
Her
bers
(19
89)
(75)
un
publ
da
ta c
ited
in
Her
bers
(19
89)
(76)
Her
bers
(19
91)
(77)
Her
bers
amp G
riec
o (1
994)
(7
8)H
erbe
rs amp
Tu
cker
(19
86)
(79)
Hof
fman
n (
1998
) (8
0) H
oumllld
oble
r (1
983)
(81
) H
oumllld
oble
r (1
984)
(82
) H
oumllld
oble
r amp
Lu
msd
en (
1980
) (8
3) H
oumllld
oble
r amp
Moumlg
lich
(19
80)
(84)
Houmll
ldob
ler
amp W
ilso
n (
1977
) (8
5) H
oumllld
oble
r amp
Wil
son
(19
86)
(86)
Houmll
ldob
ler
amp W
ilso
n (
1990
) (8
7) H
olw
ay amp
Cas
e (2
000)
(88
) Ic
hin
ose
(198
7) (
89)
Ito
Hig
ash
iamp
Mae
ta (
1988
) (9
0) J
anze
n (
1973
) (9
1) J
ayas
uri
ya amp
Tra
nie
llo
(198
5) (
92)
Joh
nso
n amp
Cro
zier
(19
98)
(93)
Kan
now
ski
(195
9) (
94)
Kas
pari
amp V
argo
(19
95)
(95)
Kel
ler
(199
1)
(96)
Kel
ler
(199
3)
(97)
var
iou
s re
fere
nce
s in
Kel
ler
(199
8)
(98)
Kel
ler
amp P
asse
ra (
1990
) (9
9) K
enn
e (1
999)
(1
00)
Kle
in (
1987
) (1
01)
Klo
tz
et a
l
(1
996)
(1
02)
Le
Mas
ne
(199
4)
(103
) L
esto
n (
1978
) (1
04)
Leacutev
ieu
x amp
Dio
man
de (
1978
) (1
05)
Lev
ieu
x (1
983)
(1
06)
Lev
ings
amp T
ran
iell
o (1
981)
(1
07)
Lie
fke
et a
l
(1
998)
(10
8) L
ongi
no
(199
1) (
109)
Lon
gin
o (2
000)
(11
0) M
abel
is (
1994
) (1
11)
Mae
der
amp C
her
ix (
2001
) (1
12)
Maj
er (
1976
) (1
13)
Mas
chw
itz
amp M
oog
(200
0) (
114)
McG
lyn
n (
1999
) (1
15)
McI
ver
(199
1) (
116)
McI
ver
amp S
teen
(19
94)
(117
) M
cKey
D (
1984
) (1
18)
Mer
cier
amp D
ejea
n (
1996
) (1
19)
Mer
cier
Len
oir
amp D
ejea
n (
1994
)(1
20)
Mer
cier
et a
l
(19
96)
(121
) M
orai
s (1
994)
(12
2) N
icke
rson
et a
l
(19
75)
(123
) O
rsquoNei
l (19
88)
(124
) O
rive
l (20
00)
(125
) P
amil
o (1
991)
(12
6) P
amil
o amp
Ros
engr
en(1
983)
(12
7) P
amil
o C
rozi
er amp
Fra
ser
(198
5) (
128)
Par
trid
ge P
artr
idge
amp F
ran
ks (
1997
) (1
29)
Pas
sera
(19
93)
(130
) P
asse
ra (
1994
) (1
31)
Pas
sera
Gil
bert
amp A
ron
(200
1)
(132
) P
eder
sen
amp B
oom
sma
(199
9)
(133
) P
eete
rs (
1993
) (1
34)
Pee
ters
amp C
rew
e (1
986)
(1
35)
Pen
g C
hri
stia
n amp
Gib
b (1
998)
(1
36)
Per
fect
o (1
994)
(1
37)
Pfe
iffe
r amp
Lin
sen
mai
r (1
998)
(13
8) P
feif
fer
amp L
inse
nm
air
(200
0) (
139)
Pfe
iffe
r amp
Lin
sen
mai
r (2
001)
(14
0) P
irk
et a
l
(20
01)
(141
) P
isar
ski
amp C
zech
owsk
i (1
990)
(1
42)
Roi
sin
et a
l
(19
86)
(143
) R
osen
gren
et a
l
(19
85)
(144
) R
owe
amp B
rist
ow (
1999
) (1
45)
Ruuml
ppel
amp H
ein
ze (
1999
) (1
46)
San
ders
amp G
ordo
n (
2000
) (1
47)
Sas
aki
Sat
oh amp
Oba
ra (
1996
) (1
48)
Sav
olai
nen
amp V
epsauml
laumlin
en (
1988
) (1
49)
Sav
olai
nen
Vep
saumllauml
inen
amp D
esli
ppe
(199
6) (
150)
Sch
mid
-Hem
pel
(198
7) (
151)
Sei
fert
(20
00)
(152
) S
mit
h-G
lase
r (1
994)
(15
3) S
nyd
er amp
Her
bers
(19
91)
(154
) S
tuar
t (1
985)
(15
5) S
tuar
t (1
987)
(15
6) S
tuar
t (1
991)
(15
7) S
un
dstr
oumlm (
1989
) (1
58)
Su
nds
troumlm
(199
3a)
(159
) T
orro
ssia
n (
1960
) (1
60)
Tor
ossi
an (
1974
) (1
61)
Tra
nie
llo
(198
2) (
162)
Tra
nie
llo
(198
9) (
163)
Tra
nie
llo
amp L
evin
gs (
1986
) (1
64)
Tsu
ji (
1988
) (1
65)
Tsu
ji amp
Yam
auch
i (1
994)
(16
6) T
suji
et a
l
(19
91)
(167
) V
asco
nce
los
amp D
avid
son
(20
00)
(168
) V
epsauml
laumlin
en
et a
l
(20
00)
(169
) W
agn
er (
1997
) (1
70)
Wag
ner
(20
00)
(171
) W
alke
r amp
Sta
mps
(19
86)
(172
) W
alof
f amp
Bla
ckit
h (
1962
) (1
73)
Way
(19
54)
(174
) Ya
mau
chi
et a
l
(19
96)
(175
) Ya
mau
chi
et a
l
(20
01)
(176
) Z
akh
arov
(19
94)
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS
321
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Tab
le 1
Lis
t of
th
e an
t sp
ecie
s (H
ymen
opte
ra
For
mic
idae
) sh
owin
g (o
blig
ate
or f
acu
ltat
ive)
pol
ydom
ous
colo
nia
l st
ruct
ure
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
PO
NE
RIN
AE
Am
blyo
poni
niA
mbl
yopo
ne p
alli
pes
ET
AE
NR
F 16
1T
161
C16
1
F16
1n
Am
blyo
poni
niP
rion
opel
ta a
mab
ilis
NE
O (C
entr
al A
mer
ica)
109
TR
RF
109
T10
9N
109
NF
109
n10
9
Am
blyo
poni
niP
rion
opel
ta m
odes
taN
EO
(Cen
tral
Am
eric
a)10
9T
RR
F 10
9T
109
N10
9N
109
F10
9n
109
Ect
atom
min
iP
arap
oner
a cl
avat
aN
EO
41T
RR
FA
41
D41
F41
n 41
Odo
ntom
achi
niO
dont
omac
hus
may
iN
EO
(Sou
th A
mer
ica)
124
TR
RF
124
A12
4C
124
VS
124
F12
4n
124
Pon
erin
iH
ypop
oner
a bo
ndro
itiE
T)napaJ(
LAP
OA
174
T17
4N
174
VS
174
F17
4n
174
Pon
erin
iP
achy
cond
y la
ber
thou
diE
TH
(Sou
th A
fric
a)42
T
EM
ZT
42N
42V
S 13
4F
134
n13
4
Pon
erin
iP
achy
cond
y la
hot
tent
ota
ET
H (S
outh
Afr
ica)
42
TE
MZ
T42
N42
F
42n
Pon
erin
iP
achy
cond
yla
goel
dii
NE
O (
Sout
h A
mer
ica)
124
TR
RF
124
A12
4C
124
N12
4F
124
n12
4
PSE
UD
OM
YR
ME
CIN
AE
Pse
udom
yrm
ecin
iP
seud
omyr
mex
eje
ctus
NE
A 10
0S
TO
AA
100
N10
0
O
100
n10
0
Pse
udom
yrm
ecin
iP
seud
omyr
mex
pal
lidus
NE
A 10
0S
TO
AA
100
N10
0
O
100
n10
0
Pse
udom
yrm
ecin
iP
seud
omyr
mex
sem
inol
eN
EA
100
ST
OA
A10
0N
100
O
10
0n
100
Pse
udom
yrm
ecin
iP
seud
omyr
mex
ven
efic
aN
EO
(Cen
tral
Am
eric
a)90
TR
RF
172
A90
N90
D90
O
90n
90
Pse
udom
yrm
ecin
iT
etra
pone
ra s
p P
SW-8
0 ne
ar a
ttenu
ata
TS
)aisAtsae -htuos(
IR
OR
F 14
A14
N14
F
14n
14
MY
RM
ICIN
AE
Cat
aula
cini
Cat
aula
cus
catu
volc
us
FN
NA
FR
RT
IR
OC
atau
laci
niC
atau
lacu
s gu
inee
nsis
RT
)ac irfAla rtne
Camptse
W(H
TE
RF
1T
1N
1N
49F
1
n
1
Cat
aula
cini
Cat
aula
cus
mck
eyi
RT
)acirfAlartne
C(H
TE
RF
117
A11
7N
117
N11
7F
44n
44
Cat
aula
cini
Cat
aula
cus
mut
icus
F
NN
AF
RR
TI
RO
Cep
halo
tini
Cep
halo
tes
atra
tus
NE
O 28
TR
RF
A28
N28
D28
F28
n28
Cep
halo
tini
Cep
halo
tes
min
utus
NE
O 10
9T
RR
FA
109
N10
9
F10
9n
109
Cep
halo
tini
Cep
halo
tes
umbr
acul
atus
NE
O 10
9T
RR
FA
109
E10
9
F10
9n
109
Cre
mat
ogas
trin
iC
rem
atog
aste
r af
rica
naE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
112
F11
2n
112
Cre
mat
ogas
trin
iC
rem
atog
aste
r br
evis
pino
saN
EO
(So
uth
Am
eric
a)41
TR
RF
A41
C41
D41
F41
n41
Cre
mat
ogas
trin
iC
rem
atog
aste
r cl
ariv
entr
isE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
112
F49
112
n11
2
Cre
mat
ogas
trin
iC
rem
atog
aste
r de
pres
saE
TH
(C
entr
al A
fric
a)11
2T
RR
FA
112
C11
2D
49 1
12F
49 1
12n
112
Cre
mat
ogas
trin
iC
rem
atog
aste
r ga
bone
nsis
ET
H (
Cen
tral
Afr
ica)
50T
RR
FA
50C
50D
50F
50n
50
Cre
mat
ogas
trin
iC
rem
atog
aste
r he
lioph
ilaE
TH
(W
est A
fric
a)53
TR
OA
53A
53N
53D
53F
53n
53
Cre
mat
ogas
trin
iC
rem
atog
aste
r im
pres
saE
TH
(W
est A
fric
a)53
TR
OA
53A
53N
53D
53F
53n
53
Cre
mat
ogas
trin
iC
rem
atog
aste
r la
evis
NE
O (
Sout
h A
mer
ica)
167
TR
RF
A16
7N
167
D16
7
n16
7
Cre
mat
ogas
trin
iC
rem
atog
aste
r lim
ata
para
biot
ica
NE
O (
Sout
h A
mer
ica)
124
TR
RF
124
A41
C41
D41
F41
n41
Cre
mat
ogas
trin
iC
rem
atog
aste
r lo
ngis
pina
F
RR
T)acire
mAlartne
C(O
EN
F7
n7
Cre
mat
ogas
trin
iC
rem
atog
aste
r sc
utel
lari
sPA
L +
OR
I 117
TE
MZ
M11
7E
117
VS
117
F11
7n
117
Cre
mat
ogas
trin
iC
rem
atog
aste
r st
riat
ula
ET
H (
Cen
tral
Afr
ica)
112
TR
RF
A11
2C
112
D49
112
F11
2n
112
Lep
toth
orac
ini
Car
dioc
ondy
la e
mer
yiE
TH
+ A
ntill
a 13
0P
AN
AH
T12
9N
129
U12
9O
(uni
col)
129
n12
9
Lep
toth
orac
ini
Car
dioc
ondy
la n
uda
AA
S +
Mad
agas
car 36
130
PA
NA
H 36
T12
9N
129
U12
9O
(uni
col)
129
n12
9
Lep
toth
orac
ini
Car
dioc
ondy
la w
roug
hton
iiH
OL
+ In
dia
130
PA
NA
HT
129
N12
9U
129
O(u
nico
l)12
9n
129
Lep
toth
orac
ini
Lep
toth
orax
am
bigu
usE
TA
EN
TF
77A
142
N77
S5
F(s
ize-
dpdt
)(s
ize-
dpdt
)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)5
77
Lep
toth
orac
ini
Lep
toth
orax
cur
visp
inos
usE
TA
EN
TF
5 7
7A
154
N15
4S
5F
5 1
54 1
55y y y y y yy
Lep
toth
orac
ini
Lep
toth
orax
long
ispi
nosu
sE
TA
EN
TF
5 7
1M
78N
5 7
1S
5F
55
71
72
78
Lep
toth
orac
ini
Lep
toth
orax
nyl
ande
riE
T) yn a
mreG(
LAP
TF
68T
68N
68N
68F
6868
Lep
toth
orac
ini
Lep
toth
orax
pill
agen
sE
T)
AS
Un re tsae -htr o
N (A
EN
TF
75T
73
S73
F73
73
Lep
toth
orac
ini
Lep
toth
orax
tube
roin
terr
rupt
usPA
L (
Eur
ope)
128
TE
OA
128
T12
8C
N12
8F
9912
8
Lep
toth
orac
ini
Pro
tom
ogna
thus
a
mer
ican
usE
T)
AS
Unr etsae-ht ro
N( ( ( (
AE
NT
F54
60
T54
60
NS
54 6
0F
54n
54 6
0
a
b b
c
Myr
mic
arii
niM
yrm
icar
ia e
umen
oide
sE
TH
W
est amp
Cen
tral
Afr
ica)
99 1
05T
RO
AT
99C
99D
99F
99n
99
Myr
mic
arii
niM
yrm
icar
ia o
paci
vent
ris
ET
H
Cen
tral
amp S
outh
Afr
ica)
99T
RO
AT
99C
(gal
leri
es)
99D
99F
99n
99
Myr
mic
ini
Myr
mic
a pu
ncti
vent
ris
ET
AE
NT
F 6
T6
N15
3 4
5 6
N6
F45
153
45
73
6
Myr
mic
ini
Myr
mic
a ru
gino
dis
(mic
rogy
na f
orm
)PA
L
Eur
ope)
168
TE
TF
168
T16
8
N16
8F
168
M
yrm
icin
iM
yrm
ica
sulc
inod
isA
OE
T)eporu
E(L
APT
132
C13
2N
Fn
132
Och
etom
yrm
ecin
iW
asm
anni
a au
ropu
ncta
taH
OL
+ C
amer
oon
(int
rodu
ced
rang
e)13
0PA
NA
HT
129
N12
9U
129
O(u
nico
l)12
9n
129
Associa
ted
gyny
Colony
size
P13
3 o
r M
161
1
161
133
87
109
133
3
85 1
09 1
33
124
4
124
100
2
100
100
2
100
100
2
100
90 9
5
8 90
84
14
5 14
1
6 1
44
3 44
28
5 28
109
2
109
109
112
41 112
51
53
6 53
41
M10
9
M
41 1
33
4 94
133
87
M12
4
4 12
4M
174
2
174
P42
134
3
94W
42
1 42
W M P P M P M M1
P NL
NL
4
M M 3
3
3
3
8
53
M M
167
2
167
M P
112
3
112
129
129
2
36
129
77
1 15
6 1
29
154
155
286
94
154
155
77 7
8
2 73
78
94
97 9
5
128
2
128
54 6
0
2 54
99
6 94
105
99
7 99
NL
NL
45
2 94
95
4 94
132
2
94
97 1
29
4 94
97 1
17
M P P P P N
LP
M
NL
M P P P P P
322
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Phe
idol
ini
Aph
aeno
gast
er c
ocke
rell
iN
EA
(So
uthw
este
rn U
SA)14
6ST
MZ
T14
6E
146
D14
6F
146
n14
6
Phe
idol
ini
Mes
sor
barb
arus
PAL
(M
edit
erre
an z
one)
2 1
9E
RM
ZT
2 1
9C
(gal
leri
es)
2 1
9D
2F
(siz
e-dp
dt)
F (s
ize-
dpdt
)
2
Phe
idol
ini
Mes
sor
cape
nsis
ET
H (
Sout
h A
fric
a) 4
3T
EO
AT
43E
43D
43F
43n
43
Phe
idol
ini
Mes
sor
was
sman
ni
TA
OR
E)eporu
Enaenarretide
M(L
APC
66
F66
P
heid
olin
iP
heid
ole
anas
tasi
iN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
D10
9F
109
n10
9
Phe
idol
ini
Phe
idol
e ar
ieta
nsN
EO
(C
entr
al A
mer
ica)
109
TR
RF
M10
9N
109
F
109
n10
9
Phe
idol
ini
Phe
idol
e de
sert
orum
nF
DE
TZ
MR
E)
ASU
htuos(A
EN
Phe
idol
ini
Phe
idol
e m
egac
epha
laE
TH
NE
O A
US
79 1
30PA
NA
H 13
0T
129
79
N12
9U
79 1
29O
(un
icol
)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
79 1
29n
79 1
29
Phe
idol
ini
Phe
idol
e pa
llid
ula
PAL
(M
edit
erra
nean
zon
e) 55
ER
MZ
55T
55N
55N
55F
55n y
55
Phe
idol
ini
Pri
stom
yrm
ex p
unge
nsPA
L (
Japa
n)16
4T
ET
F 16
4T
164
N16
4N
164
F16
416
4
Phe
idol
ini (
)P
rocr
ypto
ceru
s la
eviv
entr
isN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
F
10
9n
109
Phe
idol
ini (
)P
rocr
ypto
ceru
s m
ayri
NE
O (
Cen
tral
amp S
outh
Am
eric
a)10
9T
RR
FA
109
N10
9
F
109
n10
9
Sol
enop
sidi
niM
onom
oriu
m d
estr
ucto
rH
AN
AP) ]egnar
evitan [aidnI (
IR
OT
129
N12
9U
129
129
n12
9
Sol
enop
sidi
niM
onom
oriu
m f
lori
cola
HA
NAP
)]egnarevitan [
aid nI(I
RO
T12
9N
129
U12
912
9n
129
Sol
enop
sidi
niM
onom
oriu
m m
inim
umN
EA
(N
orth
US
A) 16
2T
EO
AT
162
E16
2N
162
F16
2n
162
Sol
enop
sidi
niM
onom
oriu
m p
hara
onis
WW
129
130
PAN
AH
T12
9N
129
U12
9O
n12
9
Sol
enop
sidi
niSo
leno
psis
gem
inat
aN
EW
(C
entr
al A
mer
ica
[nat
ive
rang
e])
136
ER
OA
T9
136
E13
6D
136
F13
6n
136
Sol
enop
sidi
niSo
leno
psis
invi
cta
NE
A(i
ntro
duce
d ra
nge)
86
TE
OA
T9
D
F
Tet
ram
orii
niT
etra
mor
ium
acu
leat
um e
FR
RT
HT
EA
50 1
12C
50 1
12D
49 5
0 1
12F
49n
50
Tet
ram
orii
niT
etra
mor
ium
afr
ican
umn
FD
CA
FR
RT
HT
ET
etra
mor
iini
Tet
ram
oriu
m c
aesp
itum
PAL
(Eur
ope)
148
86
TE
TF
148
T14
8
N14
8F
148
n14
8
AN
EU
RE
TIN
AE
Ane
uret
us s
imon
iTS
)yle visu lc xeakna
LirS(
IR
OR
F 91
T91
N91
N91
F91
D
OL
ICH
OD
ER
INA
ED
olic
hode
rini
Dol
icho
deru
s qu
adri
punc
tatu
sPA
L (
Fran
ce)15
9T
ET
FA
159
N15
9N
O15
9 1
60n
Dol
icho
deri
niD
olic
hode
rus
f b
iden
sN
EO
(Bra
zil)
52 6
8T
RR
FA
52 1
03C
103
D
103
F10
3n
103
Dol
icho
deri
niL
iom
etop
um a
picu
latu
mn
FD
CT
ZM
RT
)A
SUtse
W(A
EN
Tap
inom
ini
Azt
eca
alfa
riN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
char
tifex
spi
riti
NE
O 52
TR
RF
A52
CD
Fn
Tap
inom
ini
Azt
eca
coer
ulei
penn
isN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
para
ensi
s bo
ndar
iN
EO
52T
RR
FA
52C
DF
nT
apin
omin
iA
ztec
a cf
lan
ugin
osa
NE
O (
Bra
zil)
121
TR
RF
A12
1C
121
D12
1F
121
n12
1
Tap
inom
ini
Azt
eca
ovat
icep
sN
EO
(B
razi
l)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
cf t
raili
NE
O (
Peru
)40
TR
RF
A40
C
40N
40F
40n
40
Tap
inom
ini
Azt
eca
trig
ona
NE
O (
Pana
ma)
3T
RR
F 3
A3
C3
D3
O
3n
3
Tap
inom
ini
Dor
ymy r
mex
g h in
sana
nF
DE
TA
OE
TA
EN
Tap
inom
ini
Irid
omyr
mex
n
itid
icep
sA
US
(Aus
tral
ia)
32E
RO
AT
32E
32D
32F
32n
32
Tap
inom
ini
Irid
omyr
mex
pur
pure
usA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
san
guin
eus
AU
S (A
ustr
alia
)11
5E
RO
A41
T11
5C
115
D41
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
vir
idia
eneu
sA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n y
115
Tap
inom
ini
Lin
epith
ema
hum
ileW
W (
intr
oduc
ed r
ange
) 87
130
PAN
AH
T12
9N
129
U12
912
987
Tap
inom
ini
Tap
inom
a m
elan
ocep
halu
mW
W 15
86
PAN
AH
130
T15
129
NU
15 1
2915
129
n15
129
Tap
inom
ini
Tap
inom
a m
inut
umR
E)ailartsu
A(S
UA
TF
76T
76N
76
F76
T
apin
omin
iT
echn
omyr
mex
alb
ipes
PA
L
(Jap
an) 1
65 1
66T
ET
FA
165
N16
5N
16
6F
166
n16
5
d
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
FO
RM
ICIN
AE
Cam
pono
tini
Cam
pono
tus
abdo
min
alis
flo
rida
nus
NE
A (
Flor
ida)
101
STM
ZT
101
E10
1D
101
F10
1n
101
Cam
pono
tini
Cam
opno
tus
brut
usE
TH
(C
entr
al A
fric
a)12
0T
RR
FT
120
N12
0N
120
49
F12
0n
120
Cam
pono
tini
Cam
pono
tus
cing
ulat
usN
EO
52T
RR
FA
52N
52
Fn
Cam
pono
tini
Cam
pono
tus
detr
itus
ET
H (
Sout
h-W
est A
fric
a)31
ER
MZ
T31
C31
D31
F31
n
31
Cam
pono
tini
Cam
pono
tus
giga
sR
T) oenro
B(I
RO
RF
137
139
T13
7 1
38 1
39C
137
138
139
D13
913
9n
137
138
139
Cam
pono
tini
Cam
pono
tus
fem
orat
usN
EO
40 1
24T
RR
F 12
4A
41C
124
D41
F41
n41
19
66
79 1
29
55
3
83
164
7
164
109
1
109
109
129
129
97
8 9
7
136
9 5
9 9
5 9
7
7 86
112
95
6 94
86
M
4
P
P M W M M1
P P
P P P M M
M NL
91
2 91
159
3
159
103
52
52
121
40
3 40
3
5 3
32
11
5
7 94
86
115
115
87 9
7 1
29
8
P 15
129
3
15
76
3 76
165
5
166
101
120
52
31
6
31
M
137
138
139
5
139
40 4
1
M NL
M
M M
7
M P P3
P P P P P NL
P 5
5
M P P
Associa
ted
gyny
Colony
size
Cam
pono
tini
Cam
pono
tus
herc
ulan
eus
HO
L 16
8 8
6E
RT
F 16
8T
168
D
168
F16
8n
168
Cam
pono
tini
Cam
pono
tus
impr
essu
sN
EA
(Fl
orid
a)17
1T
ET
F 17
1A
171
N17
1D
171
F17
1n y
171
Cam
pono
tini
Cam
pono
tus
kius
iuen
sis
PAL
(so
uthe
rn J
apan
)89
STT
FA
89N
89
F(s
ize-
dpdt
)89
89
Cam
pono
tini
Cam
pono
tus
ligni
perd
usPA
L (
Eur
ope)
62T
ET
FT
62E
62D
62F
62n
62
Cam
pono
tini
Cam
pono
tus
mod
ocN
EA
(C
alif
orni
a)39
STT
FT
39E
39D
39F
39n
Cam
pono
tini
Cam
pono
tus
penn
sylv
anic
usn
FD
ET
AO
ET
AE
N
Cam
pono
tini
Cam
pono
tus
plan
atus
NE
A (
Flor
ida
[int
rodu
ced
ranp
e])
16ST
RF
A16
N
16
F16
n16
Cam
pono
tini
Cam
pono
tus
isp
1O
RI
(Sou
th-E
ast
Asi
a [M
alay
Arc
hipe
lago
])T
RR
FA
N57
NF
57n
57
Cam
pono
tini
Col
obop
sis
nipp
onic
usPA
L (
Japa
n)67
TE
TF
A67
N67
NF
67n
Gig
anti
opin
iG
igan
tiop
s de
stru
ctor
NE
O (
Fren
ch G
uian
a)21
TR
RF
T21
N21
NF
n21
97 171
89
62 16
57 67 21
M M M P M P M M M
6
86 1
68
2
171
2
89
4
86 9
4
94
3
16
5
57
2
21
4
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
323
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
y yFo
rmic
ini
Cat
agly
phis
alb
ican
sPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is b
icol
orPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is ib
eric
aPA
L (
Iber
ian
peni
nsul
a) 18
ER
MZ
T37
N37
D37
F37
n37
Form
icin
iF
orm
ica
aqui
loni
a (F
orm
ica
s s
tr)
PAL
(E
urop
e)(E
urop
e)
96T
ET
FT
96C
96D
29 9
6F
96n
29
Form
icin
iF
orm
ica
brun
iPA
L
24T
ET
FT
24C
(mou
nd)24
D24
F24
n24
Form
icin
iF
orm
ica
cine
rea
(Se
rvif
orm
ica
)PA
L (
Nor
th E
urop
e)34
176
TE
TF
176
T34
C
TC
(mou
nd)34
D34
F34
n34
Form
icin
iF
orm
ica
cuni
cula
ria
PAL
(Po
land
)35T
ET
FD
34F
nFo
rmic
ini
For
mic
a ex
sect
a (
Cop
tofo
rmic
a)
PAL
(N
orth
Eur
ope)
96T
ET
F 14
8 1
25 3
3T
96C
96D
96F
96n
96 9
7
Form
icin
iF
orm
ica
exse
ctoi
des
NE
A 14
4 8
6T
ET
FT
12C
12D
12F
12n
12
Form
icin
iF
orm
ica
haem
orrh
oida
lis
nF
DC
TF
TE
TA
EN
Form
icin
iF
orm
ica
imita
nsPA
L (
Rus
sia)
176
TE
TF
176
T17
6C
176
D17
6F
176
n17
6
Form
icin
iF
orm
ica
lugu
bris
(F
orm
ica
s s
tr)
PAL
(E
urop
e)96
TE
TF
T96
C96
D96
F22
23
96
n22
23
96
Form
icin
iF
orm
ica
nigr
ican
s (
For
mic
a s
str
)PA
L (
Cen
tral
Eur
ope)
25T
ET
FT
25C
25D
25F
25n
25
Form
icin
iF
orm
ica
obsc
urip
esN
EA
(C
entr
al U
SA)
26 2
7 1
16T
ET
FT
116
E11
6D
116
F11
6n
116
Form
icin
iF
orm
ica
opac
iven
tris
FT
ET
)AS
U(A
EN
T12
3
D12
3F
123
Fo
rmic
ini
For
mic
a pa
llide
fulv
a ni
tidiv
entr
isN
EA
(ea
ster
n U
SA
)152
TE
TF
T15
2C
152
F
152
n15
2
Form
icin
iF
orm
ica
para
lugu
bris
(F
orm
ica
s s
tr)
PAL
(Sw
itze
rlan
d)11
1T
ET
FT
111
C11
1D
111
n y
111
Form
icin
iF
orm
ica
perp
ilosa
NE
A 63
170
STT
FT
63 1
70C
63D
169
F63
169
Form
icin
iF
orm
ica
podz
olic
aE
TA
EN
OA
149
TC
N14
9F
nFo
rmic
ini
For
mic
a po
lyct
ena
(F
orm
ica
s s
tr)
PAL
(E
urop
e)8
22
96
TE
TF
8 9
6 1
48T
8 9
6 1
41C
8 9
6 1
41D
22 9
6 1
41F
8 9
6 1
41n
8 1
41
Form
icin
iF
orm
ica
prat
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
25 9
6T
ET
FT
25 9
6C
25 9
6D
25 9
6 1
40F
25 9
6 1
40n
25
Form
icin
iF
orm
ica
pres
sila
bris
(C
opto
form
ica
)PA
L (
Nor
th E
urop
e)12
6 9
6T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
rufa
(F
orm
ica
s s
tr)
PAL
96T
ET
F 14
8T
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
sang
uine
a (R
aptif
orm
ica
)PA
L (
Finl
and)
96T
ET
FT
96C
96S
35 9
6F
96n
96
Form
icin
iF
orm
ica
tran
skau
kasi
ca (
Serv
ifor
mic
a)
PAL
(Fi
nlan
d)96
TE
TF
T96
C96
D96
F96
n y
96
Form
icin
iF
orm
ica
trun
coru
m
(For
mic
a s
str
)PA
L (
Eur
ope)
157
96
TE
TF
148
T96
143
158
C96
157
D96
F96
157
96 1
57
Form
icin
iF
orm
ica
ulke
i (F
orm
ica
s s
tr)
FT
ET
AE
NT
123
D
123
F12
3
Form
icin
iF
orm
ica
ural
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
96T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
yess
ensi
s (
For
mic
a s
str
)PA
L (
Japa
n C
orea
)96T
ET
FT
96C
96D
96F
96n
96
Form
icin
i
Pro
form
ica
long
iset
aPA
L (
Spai
n)58
ER
MZ
T58
C58
D58
F58
n58
Form
icin
iC
ampo
noti
niP
olyr
hach
is a
rach
neR
RT
)aisen odnI(I
RO
FA
107
N10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
ellic
osa
FR
RT
)aisenodnI(I
RO
T10
7C
107
D10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
icol
orF
RR
T)aisenodnI(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is d
ives
PAL
(Ja
pan)
+ O
RI
Mal
aysi
a86
TR
RF
A37
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
achi
s do
ddi
AU
S (
Aus
tral
ia)92
TR
RF
92A
92C
92V
S 92
F92
Form
icin
iC
ampo
noti
niP
olyr
hach
is f
urca
taF
RR
T)aisenodnI(
IR
OA
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is il
laud
ata
FR
RT
)aknaLirS (
IR
OT
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is la
bori
osa
ET
H (
Cam
eroo
n)11
9 1
20T
RR
FT
119
120
C11
9 1
20N
49 1
19 1
20F
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
oest
aPA
L (
Japa
n)14
7T
ET
FA
147
N14
7N
147
F14
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
uell
eri
FR
RT
)ai sya laM(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is n
igro
pilo
saF
RR
T)ai sen odnI (
IR
OA
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is p
roxi
ma
FR
RT
)ai sen odnI(I
RO
T10
7C
107
N10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is s
chel
leri
chae
FR
RT
)aisyalaM(
IR
OA
107
N10
7N
F10
7
Las
iini
Las
ius
alie
nus
PAL
(E
urop
e)74
72
73
86
TE
TF
T73
N73
DF
73
Las
iini
Las
ius
flav
usPA
L (
Eng
land
)172
TE
TF
172
T17
2C
172
D17
2F
172
n10
7
n10
7
n10
7
n10
7
n92
n10
7
n10
7
n11
9
n14
7
n10
7
n10
7
n10
7
n y
107
73
n17
2
150
150
37
M M M P29
96
24 96 1
25
12 176
22 2
3 9
5 9
6
25 95 111 149
8 9
5 9
6 9
7 1
41
25 9
5 9
6
96 95 9
6
96 9
7
96 9
7
P P P P96
157
96 96 58 107
107
107
107
92 107
107 147
107
107
107
107
95 9
7
95 9
7
P P P P P P P M P P P P P P P P M M M M M M M M
3
150
4
150
4
37
4
24
3
7
86 9
4
7
22 9
4
5
86 9
4
6
149
8
22 1
48
8
94 1
48
5
148
8
23 9
4
4
58
4
107
6
107
3
107
7
86 9
4 1
07
5
107
2
107
118
119
2
147
1
107
4
107
4
107
5
107
5
172
4 4 4N
LM M M M M M
Las
iini
Las
ius
min
utus
nF
DC
TA
OE
TA
EN
Las
iini
Las
ius
negl
ectu
sPA
L (
Wes
t Asi
a [n
ativ
e] E
urop
e [i
ntro
duce
d]) 15
1P
AN
AH
129
151
T10
129
N12
9U
10 1
29F
10n y
129
Las
iini
Las
ius
neon
iger
NE
A (
Nor
th-E
aste
rn U
SA
) 163
TE
OA
163
162
T73
E16
2D
162
163
F73
73 1
62
Las
iini
Las
ius
saka
gam
iN
EA
(Ja
pan)
175
TE
OA
T17
5E
175
DF
175
n17
5
Las
iini
Pse
udol
asiu
s sp
1n
ON
CM
FR
RT
L
asii
niP
seud
olas
ius
sp 2
F
NC
F
RR
T
Las
iini
Pse
udol
asiu
s sp
3
FN
C
FR
RT
10 1
29
16
3
95
175
P M P P2
P2
P2
P3
324
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Oec
ophy
llini
Oec
ophy
lla lo
ngin
oda
RT
HT
ER
F 82
173
A41
82
173
C41
D41
49
O49
F 41
154
n41
Oec
ophy
llini
Oec
ophy
lla s
mar
agdi
naA
US
(A
ustr
alia
) 80 1
35E
RR
FA
41 8
0 1
35C
41 8
0D
80O
41 1
35n
41 8
0
Pla
giol
epid
ini
Ano
plol
epis
long
ipes
WW
130
PA
NA
HT
129
N12
9U
129
129
n y
129
Pla
giol
epid
ini
Pla
giol
epis
pyg
mea
PAL
(Fr
ance
) 131
TE
OA
T13
1N
131
VS
131
F13
1
Pre
nole
pidi
niP
arat
rech
ina
bour
boni
caH
AN
AP
WW
T12
9N
129
U12
912
9n
129
Pre
nole
pidi
niP
arat
rech
ina
flav
ipes
HO
L (
Eas
t Asi
a [n
ativ
e] U
SA
(int
rodu
ced)
88P
AN
AH
T88
Pre
nole
pidi
niP
arat
rech
ina
long
icor
nis
WW
130
PA
NA
HT
129
N12
9U
129
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
129
n12
9
TE
RM
ITID
AE
(Is
opte
ra)
Nas
utit
erm
itina
eN
asut
iterm
es c
orni
ger
NE
O
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es n
igri
ceps
NE
O (
Pana
ma)
(Pan
ama)
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es p
rinc
eps
AU
S (
New
Gui
nea)
142
TR
RF
A14
2C
142
F
142
n14
2
Ret
icul
iter
mit
inae
Ret
icul
iterm
es f
lavi
pes
NE
A (
USA
) 13
R
FA
13C
142
F
13n
13
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
41
7 86
94
41 8
0
12
9
6 13
0
131
2
131
129
88
12
9
4 13
0
M P P P P P14
2
M M
Associa
ted
gyny
Colony
size
a
Syn
onym
of
P a
nti
llan
a
b
syn
onym
of
Oph
talm
opon
e
c
syn
onym
of
Har
pago
xen
us
d
syn
onym
of
M
sem
iru
fus
e
syn
onym
of
Mac
rom
isch
oid
es a
cule
atu
s
f
syn
onym
of
Hyp
ocli
nea
g
syn
onym
of
Con
omyr
ma
h
now
com
bin
ed a
s
An
onyc
hom
yrm
a n
itid
icep
s
i
syn
onym
of
Col
obop
sis
A
ll r
efer
ence
s li
sted
are
in
corp
orat
ed i
n t
he
bibl
iogr
aph
y of
th
e ar
ticl
e (
1) A
ckon
or (
1981
) 98
3)
(2)
Aco
sta
Lop
ez amp
Ser
ran
o (1
995)
(3
) A
dam
s (1
990)
99
4)
(4)
Ada
ms
amp L
evin
gs (1
987)
(5)
All
oway
et a
l
(19
82)
(6) B
ansc
hba
ch
et a
l
(19
97)
(7) B
enzi
ng
(199
1) (
8) B
eye
et a
l
(19
97)
(9) B
hat
kar
amp V
inso
n (1
987)
(10
) Boo
msm
a
et a
l
(19
90)
(11)
Bra
un
Pee
ters
amp H
oumllld
oble
r (1
994)
(12
) B
rist
ow
et a
l
(19
92)
(13)
Bu
lmer
et a
l
(20
01)
(14)
Bu
sch
inge
r
et a
l
(19
94)
(15)
Bu
stos
amp C
her
ix (
1998
)(1
6) C
arli
n R
eeve
amp C
over
(19
93)
(17)
Cer
da amp
Ret
ana
(199
2) (
18)
Cer
da
et a
l
(19
94)
(19)
Cer
dan
amp P
rovo
st (
1990
) (2
0) C
eust
ers
(197
9) (
21)
Ch
agn
e B
eugn
onamp
Dej
ean
(20
00)
(22)
Ch
erix
(19
86)
(23)
Ch
erix
(19
87)
(24)
Ch
erix
amp M
adda
len
a-F
elle
r (1
987)
(25
) C
olli
ngw
ood
(198
7) (
26)
Con
way
(19
96)
(27)
Con
way
(19
97)
(28)
Cor
n (
1980
) (2
9) C
osen
s amp
Tou
ssai
nt
(198
5)
(30)
Cro
zier
P
amil
o amp
Cro
zier
(19
84)
(31)
Cu
rtis
(19
85)
(32)
Cu
shm
an
Ras
hbr
ook
amp B
eatt
ie (
1994
) (3
3)C
zech
owsk
i (1
990)
(3
4) C
zech
owsk
i (1
999)
(3
5) C
zech
owsk
i amp
Rot
kiew
icz
(199
4)
(36)
Cze
chow
ski
amp Y
amau
chi
(199
7)
(37)
Dah
bi (
1997
) (3
8) D
ahbi
amp L
enoi
r(1
998a
) (3
9) D
avid
amp W
ood
(198
0) (
40)
Dav
idso
n (
1988
) (4
1) D
avid
son
(19
97)
(42)
Dea
n (
1989
) (4
3) D
ean
amp Y
eato
n (
1993
) (4
4) D
ebou
t
et a
l
(20
03)
(45)
DeH
eer
Bac
kus
amp H
erbe
rs (
2001
) (4
6) D
ejea
n amp
Feacuten
eacuteron
(19
93)
(47)
Dej
ean
amp L
ach
aud
(199
4)
(48)
Dej
ean
et a
l
(1
993)
(4
9) D
ejea
n
et a
l
(1
994)
(5
0) D
ejea
n
Dji
eto-
Lor
don
amp D
ura
nd
(199
7) (
51)
Dej
ean
et a
l
(20
00)
(52)
Del
abie
Ben
ton
amp d
e M
edei
ros
(199
1) (
53)
Del
age-
Dar
chen
(19
74)
(54)
Del
Rio
Pes
ado
amp A
llow
ay (
1983
)(5
5) D
etra
in (
1990
) (5
6) E
lmes
(19
87)
(57)
Fed
erle
M
asch
wit
z amp
Fia
la (
1998
) (5
8) F
ern
ande
z-E
scu
dero
et a
l
(2
001)
(5
9) F
letc
her
et a
l
(1
980)
(6
0) F
oitz
ik amp
Her
bers
(20
01)
(61)
Fra
nco
eur
amp P
eacutepin
(19
78)
(62)
Gad
au
et a
l
(19
98)
(63)
Ger
st (
2001
) (6
4) G
iber
nau
amp D
ejea
n (
2001
) (6
5) G
reen
slad
e amp
Hal
lida
y (1
983)
(66
)H
arkn
ess
amp I
sham
(19
88)
(67)
Has
egaw
a (1
992)
(6
8) H
ein
ze
et a
l
(1
996)
(6
9) H
elm
s (1
999)
(7
0) H
elm
s
et a
l
(2
000)
(7
1) H
erbe
rs (
1986
) (7
2) H
erbe
rs (
1987
)(7
3) H
erbe
rs (
1989
) (7
4) p
ers
obs
ev
cite
d in
Her
bers
(19
89)
(75)
un
publ
da
ta c
ited
in
Her
bers
(19
89)
(76)
Her
bers
(19
91)
(77)
Her
bers
amp G
riec
o (1
994)
(7
8)H
erbe
rs amp
Tu
cker
(19
86)
(79)
Hof
fman
n (
1998
) (8
0) H
oumllld
oble
r (1
983)
(81
) H
oumllld
oble
r (1
984)
(82
) H
oumllld
oble
r amp
Lu
msd
en (
1980
) (8
3) H
oumllld
oble
r amp
Moumlg
lich
(19
80)
(84)
Houmll
ldob
ler
amp W
ilso
n (
1977
) (8
5) H
oumllld
oble
r amp
Wil
son
(19
86)
(86)
Houmll
ldob
ler
amp W
ilso
n (
1990
) (8
7) H
olw
ay amp
Cas
e (2
000)
(88
) Ic
hin
ose
(198
7) (
89)
Ito
Hig
ash
iamp
Mae
ta (
1988
) (9
0) J
anze
n (
1973
) (9
1) J
ayas
uri
ya amp
Tra
nie
llo
(198
5) (
92)
Joh
nso
n amp
Cro
zier
(19
98)
(93)
Kan
now
ski
(195
9) (
94)
Kas
pari
amp V
argo
(19
95)
(95)
Kel
ler
(199
1)
(96)
Kel
ler
(199
3)
(97)
var
iou
s re
fere
nce
s in
Kel
ler
(199
8)
(98)
Kel
ler
amp P
asse
ra (
1990
) (9
9) K
enn
e (1
999)
(1
00)
Kle
in (
1987
) (1
01)
Klo
tz
et a
l
(1
996)
(1
02)
Le
Mas
ne
(199
4)
(103
) L
esto
n (
1978
) (1
04)
Leacutev
ieu
x amp
Dio
man
de (
1978
) (1
05)
Lev
ieu
x (1
983)
(1
06)
Lev
ings
amp T
ran
iell
o (1
981)
(1
07)
Lie
fke
et a
l
(1
998)
(10
8) L
ongi
no
(199
1) (
109)
Lon
gin
o (2
000)
(11
0) M
abel
is (
1994
) (1
11)
Mae
der
amp C
her
ix (
2001
) (1
12)
Maj
er (
1976
) (1
13)
Mas
chw
itz
amp M
oog
(200
0) (
114)
McG
lyn
n (
1999
) (1
15)
McI
ver
(199
1) (
116)
McI
ver
amp S
teen
(19
94)
(117
) M
cKey
D (
1984
) (1
18)
Mer
cier
amp D
ejea
n (
1996
) (1
19)
Mer
cier
Len
oir
amp D
ejea
n (
1994
)(1
20)
Mer
cier
et a
l
(19
96)
(121
) M
orai
s (1
994)
(12
2) N
icke
rson
et a
l
(19
75)
(123
) O
rsquoNei
l (19
88)
(124
) O
rive
l (20
00)
(125
) P
amil
o (1
991)
(12
6) P
amil
o amp
Ros
engr
en(1
983)
(12
7) P
amil
o C
rozi
er amp
Fra
ser
(198
5) (
128)
Par
trid
ge P
artr
idge
amp F
ran
ks (
1997
) (1
29)
Pas
sera
(19
93)
(130
) P
asse
ra (
1994
) (1
31)
Pas
sera
Gil
bert
amp A
ron
(200
1)
(132
) P
eder
sen
amp B
oom
sma
(199
9)
(133
) P
eete
rs (
1993
) (1
34)
Pee
ters
amp C
rew
e (1
986)
(1
35)
Pen
g C
hri
stia
n amp
Gib
b (1
998)
(1
36)
Per
fect
o (1
994)
(1
37)
Pfe
iffe
r amp
Lin
sen
mai
r (1
998)
(13
8) P
feif
fer
amp L
inse
nm
air
(200
0) (
139)
Pfe
iffe
r amp
Lin
sen
mai
r (2
001)
(14
0) P
irk
et a
l
(20
01)
(141
) P
isar
ski
amp C
zech
owsk
i (1
990)
(1
42)
Roi
sin
et a
l
(19
86)
(143
) R
osen
gren
et a
l
(19
85)
(144
) R
owe
amp B
rist
ow (
1999
) (1
45)
Ruuml
ppel
amp H
ein
ze (
1999
) (1
46)
San
ders
amp G
ordo
n (
2000
) (1
47)
Sas
aki
Sat
oh amp
Oba
ra (
1996
) (1
48)
Sav
olai
nen
amp V
epsauml
laumlin
en (
1988
) (1
49)
Sav
olai
nen
Vep
saumllauml
inen
amp D
esli
ppe
(199
6) (
150)
Sch
mid
-Hem
pel
(198
7) (
151)
Sei
fert
(20
00)
(152
) S
mit
h-G
lase
r (1
994)
(15
3) S
nyd
er amp
Her
bers
(19
91)
(154
) S
tuar
t (1
985)
(15
5) S
tuar
t (1
987)
(15
6) S
tuar
t (1
991)
(15
7) S
un
dstr
oumlm (
1989
) (1
58)
Su
nds
troumlm
(199
3a)
(159
) T
orro
ssia
n (
1960
) (1
60)
Tor
ossi
an (
1974
) (1
61)
Tra
nie
llo
(198
2) (
162)
Tra
nie
llo
(198
9) (
163)
Tra
nie
llo
amp L
evin
gs (
1986
) (1
64)
Tsu
ji (
1988
) (1
65)
Tsu
ji amp
Yam
auch
i (1
994)
(16
6) T
suji
et a
l
(19
91)
(167
) V
asco
nce
los
amp D
avid
son
(20
00)
(168
) V
epsauml
laumlin
en
et a
l
(20
00)
(169
) W
agn
er (
1997
) (1
70)
Wag
ner
(20
00)
(171
) W
alke
r amp
Sta
mps
(19
86)
(172
) W
alof
f amp
Bla
ckit
h (
1962
) (1
73)
Way
(19
54)
(174
) Ya
mau
chi
et a
l
(19
96)
(175
) Ya
mau
chi
et a
l
(20
01)
(176
) Z
akh
arov
(19
94)
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292
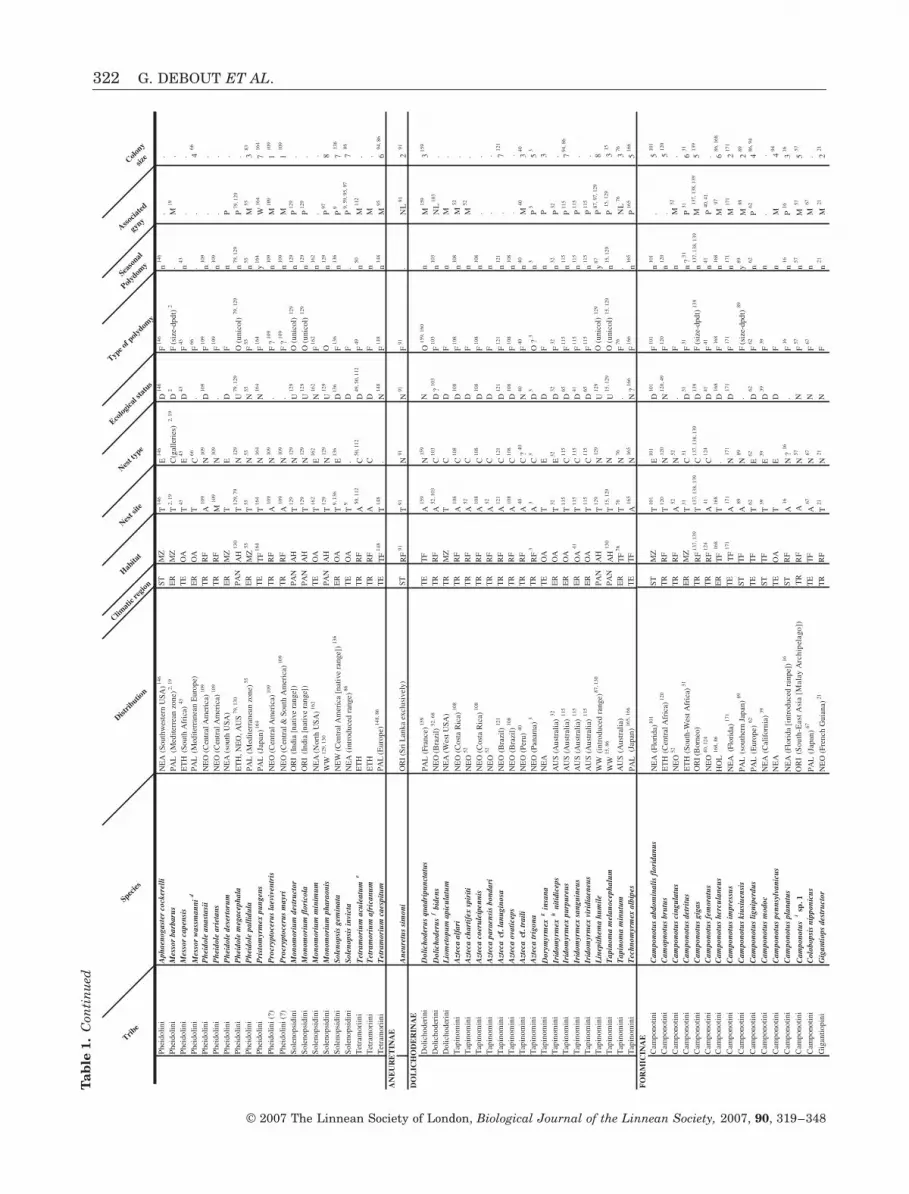
322
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Phe
idol
ini
Aph
aeno
gast
er c
ocke
rell
iN
EA
(So
uthw
este
rn U
SA)14
6ST
MZ
T14
6E
146
D14
6F
146
n14
6
Phe
idol
ini
Mes
sor
barb
arus
PAL
(M
edit
erre
an z
one)
2 1
9E
RM
ZT
2 1
9C
(gal
leri
es)
2 1
9D
2F
(siz
e-dp
dt)
F (s
ize-
dpdt
)
2
Phe
idol
ini
Mes
sor
cape
nsis
ET
H (
Sout
h A
fric
a) 4
3T
EO
AT
43E
43D
43F
43n
43
Phe
idol
ini
Mes
sor
was
sman
ni
TA
OR
E)eporu
Enaenarretide
M(L
APC
66
F66
P
heid
olin
iP
heid
ole
anas
tasi
iN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
D10
9F
109
n10
9
Phe
idol
ini
Phe
idol
e ar
ieta
nsN
EO
(C
entr
al A
mer
ica)
109
TR
RF
M10
9N
109
F
109
n10
9
Phe
idol
ini
Phe
idol
e de
sert
orum
nF
DE
TZ
MR
E)
ASU
htuos(A
EN
Phe
idol
ini
Phe
idol
e m
egac
epha
laE
TH
NE
O A
US
79 1
30PA
NA
H 13
0T
129
79
N12
9U
79 1
29O
(un
icol
)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
79 1
29n
79 1
29
Phe
idol
ini
Phe
idol
e pa
llid
ula
PAL
(M
edit
erra
nean
zon
e) 55
ER
MZ
55T
55N
55N
55F
55n y
55
Phe
idol
ini
Pri
stom
yrm
ex p
unge
nsPA
L (
Japa
n)16
4T
ET
F 16
4T
164
N16
4N
164
F16
416
4
Phe
idol
ini (
)P
rocr
ypto
ceru
s la
eviv
entr
isN
EO
(C
entr
al A
mer
ica)
109
TR
RF
A10
9N
109
F
10
9n
109
Phe
idol
ini (
)P
rocr
ypto
ceru
s m
ayri
NE
O (
Cen
tral
amp S
outh
Am
eric
a)10
9T
RR
FA
109
N10
9
F
109
n10
9
Sol
enop
sidi
niM
onom
oriu
m d
estr
ucto
rH
AN
AP) ]egnar
evitan [aidnI (
IR
OT
129
N12
9U
129
129
n12
9
Sol
enop
sidi
niM
onom
oriu
m f
lori
cola
HA
NAP
)]egnarevitan [
aid nI(I
RO
T12
9N
129
U12
912
9n
129
Sol
enop
sidi
niM
onom
oriu
m m
inim
umN
EA
(N
orth
US
A) 16
2T
EO
AT
162
E16
2N
162
F16
2n
162
Sol
enop
sidi
niM
onom
oriu
m p
hara
onis
WW
129
130
PAN
AH
T12
9N
129
U12
9O
n12
9
Sol
enop
sidi
niSo
leno
psis
gem
inat
aN
EW
(C
entr
al A
mer
ica
[nat
ive
rang
e])
136
ER
OA
T9
136
E13
6D
136
F13
6n
136
Sol
enop
sidi
niSo
leno
psis
invi
cta
NE
A(i
ntro
duce
d ra
nge)
86
TE
OA
T9
D
F
Tet
ram
orii
niT
etra
mor
ium
acu
leat
um e
FR
RT
HT
EA
50 1
12C
50 1
12D
49 5
0 1
12F
49n
50
Tet
ram
orii
niT
etra
mor
ium
afr
ican
umn
FD
CA
FR
RT
HT
ET
etra
mor
iini
Tet
ram
oriu
m c
aesp
itum
PAL
(Eur
ope)
148
86
TE
TF
148
T14
8
N14
8F
148
n14
8
AN
EU
RE
TIN
AE
Ane
uret
us s
imon
iTS
)yle visu lc xeakna
LirS(
IR
OR
F 91
T91
N91
N91
F91
D
OL
ICH
OD
ER
INA
ED
olic
hode
rini
Dol
icho
deru
s qu
adri
punc
tatu
sPA
L (
Fran
ce)15
9T
ET
FA
159
N15
9N
O15
9 1
60n
Dol
icho
deri
niD
olic
hode
rus
f b
iden
sN
EO
(Bra
zil)
52 6
8T
RR
FA
52 1
03C
103
D
103
F10
3n
103
Dol
icho
deri
niL
iom
etop
um a
picu
latu
mn
FD
CT
ZM
RT
)A
SUtse
W(A
EN
Tap
inom
ini
Azt
eca
alfa
riN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
char
tifex
spi
riti
NE
O 52
TR
RF
A52
CD
Fn
Tap
inom
ini
Azt
eca
coer
ulei
penn
isN
EO
(C
osta
Ric
a)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
para
ensi
s bo
ndar
iN
EO
52T
RR
FA
52C
DF
nT
apin
omin
iA
ztec
a cf
lan
ugin
osa
NE
O (
Bra
zil)
121
TR
RF
A12
1C
121
D12
1F
121
n12
1
Tap
inom
ini
Azt
eca
ovat
icep
sN
EO
(B
razi
l)10
8T
RR
FA
108
C10
8D
108
F10
8n
108
Tap
inom
ini
Azt
eca
cf t
raili
NE
O (
Peru
)40
TR
RF
A40
C
40N
40F
40n
40
Tap
inom
ini
Azt
eca
trig
ona
NE
O (
Pana
ma)
3T
RR
F 3
A3
C3
D3
O
3n
3
Tap
inom
ini
Dor
ymy r
mex
g h in
sana
nF
DE
TA
OE
TA
EN
Tap
inom
ini
Irid
omyr
mex
n
itid
icep
sA
US
(Aus
tral
ia)
32E
RO
AT
32E
32D
32F
32n
32
Tap
inom
ini
Irid
omyr
mex
pur
pure
usA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
san
guin
eus
AU
S (A
ustr
alia
)11
5E
RO
A41
T11
5C
115
D41
F11
5n
115
Tap
inom
ini
Irid
omyr
mex
vir
idia
eneu
sA
US
(Aus
tral
ia)
115
ER
OA
T11
5C
115
D65
F11
5n y
115
Tap
inom
ini
Lin
epith
ema
hum
ileW
W (
intr
oduc
ed r
ange
) 87
130
PAN
AH
T12
9N
129
U12
912
987
Tap
inom
ini
Tap
inom
a m
elan
ocep
halu
mW
W 15
86
PAN
AH
130
T15
129
NU
15 1
2915
129
n15
129
Tap
inom
ini
Tap
inom
a m
inut
umR
E)ailartsu
A(S
UA
TF
76T
76N
76
F76
T
apin
omin
iT
echn
omyr
mex
alb
ipes
PA
L
(Jap
an) 1
65 1
66T
ET
FA
165
N16
5N
16
6F
166
n16
5
d
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
FO
RM
ICIN
AE
Cam
pono
tini
Cam
pono
tus
abdo
min
alis
flo
rida
nus
NE
A (
Flor
ida)
101
STM
ZT
101
E10
1D
101
F10
1n
101
Cam
pono
tini
Cam
opno
tus
brut
usE
TH
(C
entr
al A
fric
a)12
0T
RR
FT
120
N12
0N
120
49
F12
0n
120
Cam
pono
tini
Cam
pono
tus
cing
ulat
usN
EO
52T
RR
FA
52N
52
Fn
Cam
pono
tini
Cam
pono
tus
detr
itus
ET
H (
Sout
h-W
est A
fric
a)31
ER
MZ
T31
C31
D31
F31
n
31
Cam
pono
tini
Cam
pono
tus
giga
sR
T) oenro
B(I
RO
RF
137
139
T13
7 1
38 1
39C
137
138
139
D13
913
9n
137
138
139
Cam
pono
tini
Cam
pono
tus
fem
orat
usN
EO
40 1
24T
RR
F 12
4A
41C
124
D41
F41
n41
19
66
79 1
29
55
3
83
164
7
164
109
1
109
109
129
129
97
8 9
7
136
9 5
9 9
5 9
7
7 86
112
95
6 94
86
M
4
P
P M W M M1
P P
P P P M M
M NL
91
2 91
159
3
159
103
52
52
121
40
3 40
3
5 3
32
11
5
7 94
86
115
115
87 9
7 1
29
8
P 15
129
3
15
76
3 76
165
5
166
101
120
52
31
6
31
M
137
138
139
5
139
40 4
1
M NL
M
M M
7
M P P3
P P P P P NL
P 5
5
M P P
Associa
ted
gyny
Colony
size
Cam
pono
tini
Cam
pono
tus
herc
ulan
eus
HO
L 16
8 8
6E
RT
F 16
8T
168
D
168
F16
8n
168
Cam
pono
tini
Cam
pono
tus
impr
essu
sN
EA
(Fl
orid
a)17
1T
ET
F 17
1A
171
N17
1D
171
F17
1n y
171
Cam
pono
tini
Cam
pono
tus
kius
iuen
sis
PAL
(so
uthe
rn J
apan
)89
STT
FA
89N
89
F(s
ize-
dpdt
)89
89
Cam
pono
tini
Cam
pono
tus
ligni
perd
usPA
L (
Eur
ope)
62T
ET
FT
62E
62D
62F
62n
62
Cam
pono
tini
Cam
pono
tus
mod
ocN
EA
(C
alif
orni
a)39
STT
FT
39E
39D
39F
39n
Cam
pono
tini
Cam
pono
tus
penn
sylv
anic
usn
FD
ET
AO
ET
AE
N
Cam
pono
tini
Cam
pono
tus
plan
atus
NE
A (
Flor
ida
[int
rodu
ced
ranp
e])
16ST
RF
A16
N
16
F16
n16
Cam
pono
tini
Cam
pono
tus
isp
1O
RI
(Sou
th-E
ast
Asi
a [M
alay
Arc
hipe
lago
])T
RR
FA
N57
NF
57n
57
Cam
pono
tini
Col
obop
sis
nipp
onic
usPA
L (
Japa
n)67
TE
TF
A67
N67
NF
67n
Gig
anti
opin
iG
igan
tiop
s de
stru
ctor
NE
O (
Fren
ch G
uian
a)21
TR
RF
T21
N21
NF
n21
97 171
89
62 16
57 67 21
M M M P M P M M M
6
86 1
68
2
171
2
89
4
86 9
4
94
3
16
5
57
2
21
4
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
323
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
y yFo
rmic
ini
Cat
agly
phis
alb
ican
sPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is b
icol
orPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is ib
eric
aPA
L (
Iber
ian
peni
nsul
a) 18
ER
MZ
T37
N37
D37
F37
n37
Form
icin
iF
orm
ica
aqui
loni
a (F
orm
ica
s s
tr)
PAL
(E
urop
e)(E
urop
e)
96T
ET
FT
96C
96D
29 9
6F
96n
29
Form
icin
iF
orm
ica
brun
iPA
L
24T
ET
FT
24C
(mou
nd)24
D24
F24
n24
Form
icin
iF
orm
ica
cine
rea
(Se
rvif
orm
ica
)PA
L (
Nor
th E
urop
e)34
176
TE
TF
176
T34
C
TC
(mou
nd)34
D34
F34
n34
Form
icin
iF
orm
ica
cuni
cula
ria
PAL
(Po
land
)35T
ET
FD
34F
nFo
rmic
ini
For
mic
a ex
sect
a (
Cop
tofo
rmic
a)
PAL
(N
orth
Eur
ope)
96T
ET
F 14
8 1
25 3
3T
96C
96D
96F
96n
96 9
7
Form
icin
iF
orm
ica
exse
ctoi
des
NE
A 14
4 8
6T
ET
FT
12C
12D
12F
12n
12
Form
icin
iF
orm
ica
haem
orrh
oida
lis
nF
DC
TF
TE
TA
EN
Form
icin
iF
orm
ica
imita
nsPA
L (
Rus
sia)
176
TE
TF
176
T17
6C
176
D17
6F
176
n17
6
Form
icin
iF
orm
ica
lugu
bris
(F
orm
ica
s s
tr)
PAL
(E
urop
e)96
TE
TF
T96
C96
D96
F22
23
96
n22
23
96
Form
icin
iF
orm
ica
nigr
ican
s (
For
mic
a s
str
)PA
L (
Cen
tral
Eur
ope)
25T
ET
FT
25C
25D
25F
25n
25
Form
icin
iF
orm
ica
obsc
urip
esN
EA
(C
entr
al U
SA)
26 2
7 1
16T
ET
FT
116
E11
6D
116
F11
6n
116
Form
icin
iF
orm
ica
opac
iven
tris
FT
ET
)AS
U(A
EN
T12
3
D12
3F
123
Fo
rmic
ini
For
mic
a pa
llide
fulv
a ni
tidiv
entr
isN
EA
(ea
ster
n U
SA
)152
TE
TF
T15
2C
152
F
152
n15
2
Form
icin
iF
orm
ica
para
lugu
bris
(F
orm
ica
s s
tr)
PAL
(Sw
itze
rlan
d)11
1T
ET
FT
111
C11
1D
111
n y
111
Form
icin
iF
orm
ica
perp
ilosa
NE
A 63
170
STT
FT
63 1
70C
63D
169
F63
169
Form
icin
iF
orm
ica
podz
olic
aE
TA
EN
OA
149
TC
N14
9F
nFo
rmic
ini
For
mic
a po
lyct
ena
(F
orm
ica
s s
tr)
PAL
(E
urop
e)8
22
96
TE
TF
8 9
6 1
48T
8 9
6 1
41C
8 9
6 1
41D
22 9
6 1
41F
8 9
6 1
41n
8 1
41
Form
icin
iF
orm
ica
prat
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
25 9
6T
ET
FT
25 9
6C
25 9
6D
25 9
6 1
40F
25 9
6 1
40n
25
Form
icin
iF
orm
ica
pres
sila
bris
(C
opto
form
ica
)PA
L (
Nor
th E
urop
e)12
6 9
6T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
rufa
(F
orm
ica
s s
tr)
PAL
96T
ET
F 14
8T
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
sang
uine
a (R
aptif
orm
ica
)PA
L (
Finl
and)
96T
ET
FT
96C
96S
35 9
6F
96n
96
Form
icin
iF
orm
ica
tran
skau
kasi
ca (
Serv
ifor
mic
a)
PAL
(Fi
nlan
d)96
TE
TF
T96
C96
D96
F96
n y
96
Form
icin
iF
orm
ica
trun
coru
m
(For
mic
a s
str
)PA
L (
Eur
ope)
157
96
TE
TF
148
T96
143
158
C96
157
D96
F96
157
96 1
57
Form
icin
iF
orm
ica
ulke
i (F
orm
ica
s s
tr)
FT
ET
AE
NT
123
D
123
F12
3
Form
icin
iF
orm
ica
ural
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
96T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
yess
ensi
s (
For
mic
a s
str
)PA
L (
Japa
n C
orea
)96T
ET
FT
96C
96D
96F
96n
96
Form
icin
i
Pro
form
ica
long
iset
aPA
L (
Spai
n)58
ER
MZ
T58
C58
D58
F58
n58
Form
icin
iC
ampo
noti
niP
olyr
hach
is a
rach
neR
RT
)aisen odnI(I
RO
FA
107
N10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
ellic
osa
FR
RT
)aisenodnI(I
RO
T10
7C
107
D10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
icol
orF
RR
T)aisenodnI(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is d
ives
PAL
(Ja
pan)
+ O
RI
Mal
aysi
a86
TR
RF
A37
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
achi
s do
ddi
AU
S (
Aus
tral
ia)92
TR
RF
92A
92C
92V
S 92
F92
Form
icin
iC
ampo
noti
niP
olyr
hach
is f
urca
taF
RR
T)aisenodnI(
IR
OA
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is il
laud
ata
FR
RT
)aknaLirS (
IR
OT
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is la
bori
osa
ET
H (
Cam
eroo
n)11
9 1
20T
RR
FT
119
120
C11
9 1
20N
49 1
19 1
20F
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
oest
aPA
L (
Japa
n)14
7T
ET
FA
147
N14
7N
147
F14
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
uell
eri
FR
RT
)ai sya laM(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is n
igro
pilo
saF
RR
T)ai sen odnI (
IR
OA
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is p
roxi
ma
FR
RT
)ai sen odnI(I
RO
T10
7C
107
N10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is s
chel
leri
chae
FR
RT
)aisyalaM(
IR
OA
107
N10
7N
F10
7
Las
iini
Las
ius
alie
nus
PAL
(E
urop
e)74
72
73
86
TE
TF
T73
N73
DF
73
Las
iini
Las
ius
flav
usPA
L (
Eng
land
)172
TE
TF
172
T17
2C
172
D17
2F
172
n10
7
n10
7
n10
7
n10
7
n92
n10
7
n10
7
n11
9
n14
7
n10
7
n10
7
n10
7
n y
107
73
n17
2
150
150
37
M M M P29
96
24 96 1
25
12 176
22 2
3 9
5 9
6
25 95 111 149
8 9
5 9
6 9
7 1
41
25 9
5 9
6
96 95 9
6
96 9
7
96 9
7
P P P P96
157
96 96 58 107
107
107
107
92 107
107 147
107
107
107
107
95 9
7
95 9
7
P P P P P P P M P P P P P P P P M M M M M M M M
3
150
4
150
4
37
4
24
3
7
86 9
4
7
22 9
4
5
86 9
4
6
149
8
22 1
48
8
94 1
48
5
148
8
23 9
4
4
58
4
107
6
107
3
107
7
86 9
4 1
07
5
107
2
107
118
119
2
147
1
107
4
107
4
107
5
107
5
172
4 4 4N
LM M M M M M
Las
iini
Las
ius
min
utus
nF
DC
TA
OE
TA
EN
Las
iini
Las
ius
negl
ectu
sPA
L (
Wes
t Asi
a [n
ativ
e] E
urop
e [i
ntro
duce
d]) 15
1P
AN
AH
129
151
T10
129
N12
9U
10 1
29F
10n y
129
Las
iini
Las
ius
neon
iger
NE
A (
Nor
th-E
aste
rn U
SA
) 163
TE
OA
163
162
T73
E16
2D
162
163
F73
73 1
62
Las
iini
Las
ius
saka
gam
iN
EA
(Ja
pan)
175
TE
OA
T17
5E
175
DF
175
n17
5
Las
iini
Pse
udol
asiu
s sp
1n
ON
CM
FR
RT
L
asii
niP
seud
olas
ius
sp 2
F
NC
F
RR
T
Las
iini
Pse
udol
asiu
s sp
3
FN
C
FR
RT
10 1
29
16
3
95
175
P M P P2
P2
P2
P3
324
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Oec
ophy
llini
Oec
ophy
lla lo
ngin
oda
RT
HT
ER
F 82
173
A41
82
173
C41
D41
49
O49
F 41
154
n41
Oec
ophy
llini
Oec
ophy
lla s
mar
agdi
naA
US
(A
ustr
alia
) 80 1
35E
RR
FA
41 8
0 1
35C
41 8
0D
80O
41 1
35n
41 8
0
Pla
giol
epid
ini
Ano
plol
epis
long
ipes
WW
130
PA
NA
HT
129
N12
9U
129
129
n y
129
Pla
giol
epid
ini
Pla
giol
epis
pyg
mea
PAL
(Fr
ance
) 131
TE
OA
T13
1N
131
VS
131
F13
1
Pre
nole
pidi
niP
arat
rech
ina
bour
boni
caH
AN
AP
WW
T12
9N
129
U12
912
9n
129
Pre
nole
pidi
niP
arat
rech
ina
flav
ipes
HO
L (
Eas
t Asi
a [n
ativ
e] U
SA
(int
rodu
ced)
88P
AN
AH
T88
Pre
nole
pidi
niP
arat
rech
ina
long
icor
nis
WW
130
PA
NA
HT
129
N12
9U
129
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
129
n12
9
TE
RM
ITID
AE
(Is
opte
ra)
Nas
utit
erm
itina
eN
asut
iterm
es c
orni
ger
NE
O
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es n
igri
ceps
NE
O (
Pana
ma)
(Pan
ama)
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es p
rinc
eps
AU
S (
New
Gui
nea)
142
TR
RF
A14
2C
142
F
142
n14
2
Ret
icul
iter
mit
inae
Ret
icul
iterm
es f
lavi
pes
NE
A (
USA
) 13
R
FA
13C
142
F
13n
13
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
41
7 86
94
41 8
0
12
9
6 13
0
131
2
131
129
88
12
9
4 13
0
M P P P P P14
2
M M
Associa
ted
gyny
Colony
size
a
Syn
onym
of
P a
nti
llan
a
b
syn
onym
of
Oph
talm
opon
e
c
syn
onym
of
Har
pago
xen
us
d
syn
onym
of
M
sem
iru
fus
e
syn
onym
of
Mac
rom
isch
oid
es a
cule
atu
s
f
syn
onym
of
Hyp
ocli
nea
g
syn
onym
of
Con
omyr
ma
h
now
com
bin
ed a
s
An
onyc
hom
yrm
a n
itid
icep
s
i
syn
onym
of
Col
obop
sis
A
ll r
efer
ence
s li
sted
are
in
corp
orat
ed i
n t
he
bibl
iogr
aph
y of
th
e ar
ticl
e (
1) A
ckon
or (
1981
) 98
3)
(2)
Aco
sta
Lop
ez amp
Ser
ran
o (1
995)
(3
) A
dam
s (1
990)
99
4)
(4)
Ada
ms
amp L
evin
gs (1
987)
(5)
All
oway
et a
l
(19
82)
(6) B
ansc
hba
ch
et a
l
(19
97)
(7) B
enzi
ng
(199
1) (
8) B
eye
et a
l
(19
97)
(9) B
hat
kar
amp V
inso
n (1
987)
(10
) Boo
msm
a
et a
l
(19
90)
(11)
Bra
un
Pee
ters
amp H
oumllld
oble
r (1
994)
(12
) B
rist
ow
et a
l
(19
92)
(13)
Bu
lmer
et a
l
(20
01)
(14)
Bu
sch
inge
r
et a
l
(19
94)
(15)
Bu
stos
amp C
her
ix (
1998
)(1
6) C
arli
n R
eeve
amp C
over
(19
93)
(17)
Cer
da amp
Ret
ana
(199
2) (
18)
Cer
da
et a
l
(19
94)
(19)
Cer
dan
amp P
rovo
st (
1990
) (2
0) C
eust
ers
(197
9) (
21)
Ch
agn
e B
eugn
onamp
Dej
ean
(20
00)
(22)
Ch
erix
(19
86)
(23)
Ch
erix
(19
87)
(24)
Ch
erix
amp M
adda
len
a-F
elle
r (1
987)
(25
) C
olli
ngw
ood
(198
7) (
26)
Con
way
(19
96)
(27)
Con
way
(19
97)
(28)
Cor
n (
1980
) (2
9) C
osen
s amp
Tou
ssai
nt
(198
5)
(30)
Cro
zier
P
amil
o amp
Cro
zier
(19
84)
(31)
Cu
rtis
(19
85)
(32)
Cu
shm
an
Ras
hbr
ook
amp B
eatt
ie (
1994
) (3
3)C
zech
owsk
i (1
990)
(3
4) C
zech
owsk
i (1
999)
(3
5) C
zech
owsk
i amp
Rot
kiew
icz
(199
4)
(36)
Cze
chow
ski
amp Y
amau
chi
(199
7)
(37)
Dah
bi (
1997
) (3
8) D
ahbi
amp L
enoi
r(1
998a
) (3
9) D
avid
amp W
ood
(198
0) (
40)
Dav
idso
n (
1988
) (4
1) D
avid
son
(19
97)
(42)
Dea
n (
1989
) (4
3) D
ean
amp Y
eato
n (
1993
) (4
4) D
ebou
t
et a
l
(20
03)
(45)
DeH
eer
Bac
kus
amp H
erbe
rs (
2001
) (4
6) D
ejea
n amp
Feacuten
eacuteron
(19
93)
(47)
Dej
ean
amp L
ach
aud
(199
4)
(48)
Dej
ean
et a
l
(1
993)
(4
9) D
ejea
n
et a
l
(1
994)
(5
0) D
ejea
n
Dji
eto-
Lor
don
amp D
ura
nd
(199
7) (
51)
Dej
ean
et a
l
(20
00)
(52)
Del
abie
Ben
ton
amp d
e M
edei
ros
(199
1) (
53)
Del
age-
Dar
chen
(19
74)
(54)
Del
Rio
Pes
ado
amp A
llow
ay (
1983
)(5
5) D
etra
in (
1990
) (5
6) E
lmes
(19
87)
(57)
Fed
erle
M
asch
wit
z amp
Fia
la (
1998
) (5
8) F
ern
ande
z-E
scu
dero
et a
l
(2
001)
(5
9) F
letc
her
et a
l
(1
980)
(6
0) F
oitz
ik amp
Her
bers
(20
01)
(61)
Fra
nco
eur
amp P
eacutepin
(19
78)
(62)
Gad
au
et a
l
(19
98)
(63)
Ger
st (
2001
) (6
4) G
iber
nau
amp D
ejea
n (
2001
) (6
5) G
reen
slad
e amp
Hal
lida
y (1
983)
(66
)H
arkn
ess
amp I
sham
(19
88)
(67)
Has
egaw
a (1
992)
(6
8) H
ein
ze
et a
l
(1
996)
(6
9) H
elm
s (1
999)
(7
0) H
elm
s
et a
l
(2
000)
(7
1) H
erbe
rs (
1986
) (7
2) H
erbe
rs (
1987
)(7
3) H
erbe
rs (
1989
) (7
4) p
ers
obs
ev
cite
d in
Her
bers
(19
89)
(75)
un
publ
da
ta c
ited
in
Her
bers
(19
89)
(76)
Her
bers
(19
91)
(77)
Her
bers
amp G
riec
o (1
994)
(7
8)H
erbe
rs amp
Tu
cker
(19
86)
(79)
Hof
fman
n (
1998
) (8
0) H
oumllld
oble
r (1
983)
(81
) H
oumllld
oble
r (1
984)
(82
) H
oumllld
oble
r amp
Lu
msd
en (
1980
) (8
3) H
oumllld
oble
r amp
Moumlg
lich
(19
80)
(84)
Houmll
ldob
ler
amp W
ilso
n (
1977
) (8
5) H
oumllld
oble
r amp
Wil
son
(19
86)
(86)
Houmll
ldob
ler
amp W
ilso
n (
1990
) (8
7) H
olw
ay amp
Cas
e (2
000)
(88
) Ic
hin
ose
(198
7) (
89)
Ito
Hig
ash
iamp
Mae
ta (
1988
) (9
0) J
anze
n (
1973
) (9
1) J
ayas
uri
ya amp
Tra
nie
llo
(198
5) (
92)
Joh
nso
n amp
Cro
zier
(19
98)
(93)
Kan
now
ski
(195
9) (
94)
Kas
pari
amp V
argo
(19
95)
(95)
Kel
ler
(199
1)
(96)
Kel
ler
(199
3)
(97)
var
iou
s re
fere
nce
s in
Kel
ler
(199
8)
(98)
Kel
ler
amp P
asse
ra (
1990
) (9
9) K
enn
e (1
999)
(1
00)
Kle
in (
1987
) (1
01)
Klo
tz
et a
l
(1
996)
(1
02)
Le
Mas
ne
(199
4)
(103
) L
esto
n (
1978
) (1
04)
Leacutev
ieu
x amp
Dio
man
de (
1978
) (1
05)
Lev
ieu
x (1
983)
(1
06)
Lev
ings
amp T
ran
iell
o (1
981)
(1
07)
Lie
fke
et a
l
(1
998)
(10
8) L
ongi
no
(199
1) (
109)
Lon
gin
o (2
000)
(11
0) M
abel
is (
1994
) (1
11)
Mae
der
amp C
her
ix (
2001
) (1
12)
Maj
er (
1976
) (1
13)
Mas
chw
itz
amp M
oog
(200
0) (
114)
McG
lyn
n (
1999
) (1
15)
McI
ver
(199
1) (
116)
McI
ver
amp S
teen
(19
94)
(117
) M
cKey
D (
1984
) (1
18)
Mer
cier
amp D
ejea
n (
1996
) (1
19)
Mer
cier
Len
oir
amp D
ejea
n (
1994
)(1
20)
Mer
cier
et a
l
(19
96)
(121
) M
orai
s (1
994)
(12
2) N
icke
rson
et a
l
(19
75)
(123
) O
rsquoNei
l (19
88)
(124
) O
rive
l (20
00)
(125
) P
amil
o (1
991)
(12
6) P
amil
o amp
Ros
engr
en(1
983)
(12
7) P
amil
o C
rozi
er amp
Fra
ser
(198
5) (
128)
Par
trid
ge P
artr
idge
amp F
ran
ks (
1997
) (1
29)
Pas
sera
(19
93)
(130
) P
asse
ra (
1994
) (1
31)
Pas
sera
Gil
bert
amp A
ron
(200
1)
(132
) P
eder
sen
amp B
oom
sma
(199
9)
(133
) P
eete
rs (
1993
) (1
34)
Pee
ters
amp C
rew
e (1
986)
(1
35)
Pen
g C
hri
stia
n amp
Gib
b (1
998)
(1
36)
Per
fect
o (1
994)
(1
37)
Pfe
iffe
r amp
Lin
sen
mai
r (1
998)
(13
8) P
feif
fer
amp L
inse
nm
air
(200
0) (
139)
Pfe
iffe
r amp
Lin
sen
mai
r (2
001)
(14
0) P
irk
et a
l
(20
01)
(141
) P
isar
ski
amp C
zech
owsk
i (1
990)
(1
42)
Roi
sin
et a
l
(19
86)
(143
) R
osen
gren
et a
l
(19
85)
(144
) R
owe
amp B
rist
ow (
1999
) (1
45)
Ruuml
ppel
amp H
ein
ze (
1999
) (1
46)
San
ders
amp G
ordo
n (
2000
) (1
47)
Sas
aki
Sat
oh amp
Oba
ra (
1996
) (1
48)
Sav
olai
nen
amp V
epsauml
laumlin
en (
1988
) (1
49)
Sav
olai
nen
Vep
saumllauml
inen
amp D
esli
ppe
(199
6) (
150)
Sch
mid
-Hem
pel
(198
7) (
151)
Sei
fert
(20
00)
(152
) S
mit
h-G
lase
r (1
994)
(15
3) S
nyd
er amp
Her
bers
(19
91)
(154
) S
tuar
t (1
985)
(15
5) S
tuar
t (1
987)
(15
6) S
tuar
t (1
991)
(15
7) S
un
dstr
oumlm (
1989
) (1
58)
Su
nds
troumlm
(199
3a)
(159
) T
orro
ssia
n (
1960
) (1
60)
Tor
ossi
an (
1974
) (1
61)
Tra
nie
llo
(198
2) (
162)
Tra
nie
llo
(198
9) (
163)
Tra
nie
llo
amp L
evin
gs (
1986
) (1
64)
Tsu
ji (
1988
) (1
65)
Tsu
ji amp
Yam
auch
i (1
994)
(16
6) T
suji
et a
l
(19
91)
(167
) V
asco
nce
los
amp D
avid
son
(20
00)
(168
) V
epsauml
laumlin
en
et a
l
(20
00)
(169
) W
agn
er (
1997
) (1
70)
Wag
ner
(20
00)
(171
) W
alke
r amp
Sta
mps
(19
86)
(172
) W
alof
f amp
Bla
ckit
h (
1962
) (1
73)
Way
(19
54)
(174
) Ya
mau
chi
et a
l
(19
96)
(175
) Ya
mau
chi
et a
l
(20
01)
(176
) Z
akh
arov
(19
94)
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS
323
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
(siz
e-dp
dt)
y yFo
rmic
ini
Cat
agly
phis
alb
ican
sPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is b
icol
orPA
L (
Nor
th A
fric
a)15
0E
RM
ZT
150
E15
0D
150
F15
015
0
Form
icin
iC
atag
lyph
is ib
eric
aPA
L (
Iber
ian
peni
nsul
a) 18
ER
MZ
T37
N37
D37
F37
n37
Form
icin
iF
orm
ica
aqui
loni
a (F
orm
ica
s s
tr)
PAL
(E
urop
e)(E
urop
e)
96T
ET
FT
96C
96D
29 9
6F
96n
29
Form
icin
iF
orm
ica
brun
iPA
L
24T
ET
FT
24C
(mou
nd)24
D24
F24
n24
Form
icin
iF
orm
ica
cine
rea
(Se
rvif
orm
ica
)PA
L (
Nor
th E
urop
e)34
176
TE
TF
176
T34
C
TC
(mou
nd)34
D34
F34
n34
Form
icin
iF
orm
ica
cuni
cula
ria
PAL
(Po
land
)35T
ET
FD
34F
nFo
rmic
ini
For
mic
a ex
sect
a (
Cop
tofo
rmic
a)
PAL
(N
orth
Eur
ope)
96T
ET
F 14
8 1
25 3
3T
96C
96D
96F
96n
96 9
7
Form
icin
iF
orm
ica
exse
ctoi
des
NE
A 14
4 8
6T
ET
FT
12C
12D
12F
12n
12
Form
icin
iF
orm
ica
haem
orrh
oida
lis
nF
DC
TF
TE
TA
EN
Form
icin
iF
orm
ica
imita
nsPA
L (
Rus
sia)
176
TE
TF
176
T17
6C
176
D17
6F
176
n17
6
Form
icin
iF
orm
ica
lugu
bris
(F
orm
ica
s s
tr)
PAL
(E
urop
e)96
TE
TF
T96
C96
D96
F22
23
96
n22
23
96
Form
icin
iF
orm
ica
nigr
ican
s (
For
mic
a s
str
)PA
L (
Cen
tral
Eur
ope)
25T
ET
FT
25C
25D
25F
25n
25
Form
icin
iF
orm
ica
obsc
urip
esN
EA
(C
entr
al U
SA)
26 2
7 1
16T
ET
FT
116
E11
6D
116
F11
6n
116
Form
icin
iF
orm
ica
opac
iven
tris
FT
ET
)AS
U(A
EN
T12
3
D12
3F
123
Fo
rmic
ini
For
mic
a pa
llide
fulv
a ni
tidiv
entr
isN
EA
(ea
ster
n U
SA
)152
TE
TF
T15
2C
152
F
152
n15
2
Form
icin
iF
orm
ica
para
lugu
bris
(F
orm
ica
s s
tr)
PAL
(Sw
itze
rlan
d)11
1T
ET
FT
111
C11
1D
111
n y
111
Form
icin
iF
orm
ica
perp
ilosa
NE
A 63
170
STT
FT
63 1
70C
63D
169
F63
169
Form
icin
iF
orm
ica
podz
olic
aE
TA
EN
OA
149
TC
N14
9F
nFo
rmic
ini
For
mic
a po
lyct
ena
(F
orm
ica
s s
tr)
PAL
(E
urop
e)8
22
96
TE
TF
8 9
6 1
48T
8 9
6 1
41C
8 9
6 1
41D
22 9
6 1
41F
8 9
6 1
41n
8 1
41
Form
icin
iF
orm
ica
prat
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
25 9
6T
ET
FT
25 9
6C
25 9
6D
25 9
6 1
40F
25 9
6 1
40n
25
Form
icin
iF
orm
ica
pres
sila
bris
(C
opto
form
ica
)PA
L (
Nor
th E
urop
e)12
6 9
6T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
rufa
(F
orm
ica
s s
tr)
PAL
96T
ET
F 14
8T
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
sang
uine
a (R
aptif
orm
ica
)PA
L (
Finl
and)
96T
ET
FT
96C
96S
35 9
6F
96n
96
Form
icin
iF
orm
ica
tran
skau
kasi
ca (
Serv
ifor
mic
a)
PAL
(Fi
nlan
d)96
TE
TF
T96
C96
D96
F96
n y
96
Form
icin
iF
orm
ica
trun
coru
m
(For
mic
a s
str
)PA
L (
Eur
ope)
157
96
TE
TF
148
T96
143
158
C96
157
D96
F96
157
96 1
57
Form
icin
iF
orm
ica
ulke
i (F
orm
ica
s s
tr)
FT
ET
AE
NT
123
D
123
F12
3
Form
icin
iF
orm
ica
ural
ensi
s (
For
mic
a s
str
)PA
L (
Eur
ope)
96T
ET
FT
96C
96D
96F
96n
96
Form
icin
iF
orm
ica
yess
ensi
s (
For
mic
a s
str
)PA
L (
Japa
n C
orea
)96T
ET
FT
96C
96D
96F
96n
96
Form
icin
i
Pro
form
ica
long
iset
aPA
L (
Spai
n)58
ER
MZ
T58
C58
D58
F58
n58
Form
icin
iC
ampo
noti
niP
olyr
hach
is a
rach
neR
RT
)aisen odnI(I
RO
FA
107
N10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
ellic
osa
FR
RT
)aisenodnI(I
RO
T10
7C
107
D10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is b
icol
orF
RR
T)aisenodnI(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is d
ives
PAL
(Ja
pan)
+ O
RI
Mal
aysi
a86
TR
RF
A37
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
achi
s do
ddi
AU
S (
Aus
tral
ia)92
TR
RF
92A
92C
92V
S 92
F92
Form
icin
iC
ampo
noti
niP
olyr
hach
is f
urca
taF
RR
T)aisenodnI(
IR
OA
107
C10
7N
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is il
laud
ata
FR
RT
)aknaLirS (
IR
OT
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is la
bori
osa
ET
H (
Cam
eroo
n)11
9 1
20T
RR
FT
119
120
C11
9 1
20N
49 1
19 1
20F
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
oest
aPA
L (
Japa
n)14
7T
ET
FA
147
N14
7N
147
F14
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is m
uell
eri
FR
RT
)ai sya laM(
IR
OA
107
C10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is n
igro
pilo
saF
RR
T)ai sen odnI (
IR
OA
107
N10
7N
107
F10
7
Form
icin
iC
ampo
noti
niP
olyr
hach
is p
roxi
ma
FR
RT
)ai sen odnI(I
RO
T10
7C
107
N10
7F
107
Form
icin
iC
ampo
noti
niP
olyr
hach
is s
chel
leri
chae
FR
RT
)aisyalaM(
IR
OA
107
N10
7N
F10
7
Las
iini
Las
ius
alie
nus
PAL
(E
urop
e)74
72
73
86
TE
TF
T73
N73
DF
73
Las
iini
Las
ius
flav
usPA
L (
Eng
land
)172
TE
TF
172
T17
2C
172
D17
2F
172
n10
7
n10
7
n10
7
n10
7
n92
n10
7
n10
7
n11
9
n14
7
n10
7
n10
7
n10
7
n y
107
73
n17
2
150
150
37
M M M P29
96
24 96 1
25
12 176
22 2
3 9
5 9
6
25 95 111 149
8 9
5 9
6 9
7 1
41
25 9
5 9
6
96 95 9
6
96 9
7
96 9
7
P P P P96
157
96 96 58 107
107
107
107
92 107
107 147
107
107
107
107
95 9
7
95 9
7
P P P P P P P M P P P P P P P P M M M M M M M M
3
150
4
150
4
37
4
24
3
7
86 9
4
7
22 9
4
5
86 9
4
6
149
8
22 1
48
8
94 1
48
5
148
8
23 9
4
4
58
4
107
6
107
3
107
7
86 9
4 1
07
5
107
2
107
118
119
2
147
1
107
4
107
4
107
5
107
5
172
4 4 4N
LM M M M M M
Las
iini
Las
ius
min
utus
nF
DC
TA
OE
TA
EN
Las
iini
Las
ius
negl
ectu
sPA
L (
Wes
t Asi
a [n
ativ
e] E
urop
e [i
ntro
duce
d]) 15
1P
AN
AH
129
151
T10
129
N12
9U
10 1
29F
10n y
129
Las
iini
Las
ius
neon
iger
NE
A (
Nor
th-E
aste
rn U
SA
) 163
TE
OA
163
162
T73
E16
2D
162
163
F73
73 1
62
Las
iini
Las
ius
saka
gam
iN
EA
(Ja
pan)
175
TE
OA
T17
5E
175
DF
175
n17
5
Las
iini
Pse
udol
asiu
s sp
1n
ON
CM
FR
RT
L
asii
niP
seud
olas
ius
sp 2
F
NC
F
RR
T
Las
iini
Pse
udol
asiu
s sp
3
FN
C
FR
RT
10 1
29
16
3
95
175
P M P P2
P2
P2
P3
324
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Oec
ophy
llini
Oec
ophy
lla lo
ngin
oda
RT
HT
ER
F 82
173
A41
82
173
C41
D41
49
O49
F 41
154
n41
Oec
ophy
llini
Oec
ophy
lla s
mar
agdi
naA
US
(A
ustr
alia
) 80 1
35E
RR
FA
41 8
0 1
35C
41 8
0D
80O
41 1
35n
41 8
0
Pla
giol
epid
ini
Ano
plol
epis
long
ipes
WW
130
PA
NA
HT
129
N12
9U
129
129
n y
129
Pla
giol
epid
ini
Pla
giol
epis
pyg
mea
PAL
(Fr
ance
) 131
TE
OA
T13
1N
131
VS
131
F13
1
Pre
nole
pidi
niP
arat
rech
ina
bour
boni
caH
AN
AP
WW
T12
9N
129
U12
912
9n
129
Pre
nole
pidi
niP
arat
rech
ina
flav
ipes
HO
L (
Eas
t Asi
a [n
ativ
e] U
SA
(int
rodu
ced)
88P
AN
AH
T88
Pre
nole
pidi
niP
arat
rech
ina
long
icor
nis
WW
130
PA
NA
HT
129
N12
9U
129
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
129
n12
9
TE
RM
ITID
AE
(Is
opte
ra)
Nas
utit
erm
itina
eN
asut
iterm
es c
orni
ger
NE
O
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es n
igri
ceps
NE
O (
Pana
ma)
(Pan
ama)
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es p
rinc
eps
AU
S (
New
Gui
nea)
142
TR
RF
A14
2C
142
F
142
n14
2
Ret
icul
iter
mit
inae
Ret
icul
iterm
es f
lavi
pes
NE
A (
USA
) 13
R
FA
13C
142
F
13n
13
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
41
7 86
94
41 8
0
12
9
6 13
0
131
2
131
129
88
12
9
4 13
0
M P P P P P14
2
M M
Associa
ted
gyny
Colony
size
a
Syn
onym
of
P a
nti
llan
a
b
syn
onym
of
Oph
talm
opon
e
c
syn
onym
of
Har
pago
xen
us
d
syn
onym
of
M
sem
iru
fus
e
syn
onym
of
Mac
rom
isch
oid
es a
cule
atu
s
f
syn
onym
of
Hyp
ocli
nea
g
syn
onym
of
Con
omyr
ma
h
now
com
bin
ed a
s
An
onyc
hom
yrm
a n
itid
icep
s
i
syn
onym
of
Col
obop
sis
A
ll r
efer
ence
s li
sted
are
in
corp
orat
ed i
n t
he
bibl
iogr
aph
y of
th
e ar
ticl
e (
1) A
ckon
or (
1981
) 98
3)
(2)
Aco
sta
Lop
ez amp
Ser
ran
o (1
995)
(3
) A
dam
s (1
990)
99
4)
(4)
Ada
ms
amp L
evin
gs (1
987)
(5)
All
oway
et a
l
(19
82)
(6) B
ansc
hba
ch
et a
l
(19
97)
(7) B
enzi
ng
(199
1) (
8) B
eye
et a
l
(19
97)
(9) B
hat
kar
amp V
inso
n (1
987)
(10
) Boo
msm
a
et a
l
(19
90)
(11)
Bra
un
Pee
ters
amp H
oumllld
oble
r (1
994)
(12
) B
rist
ow
et a
l
(19
92)
(13)
Bu
lmer
et a
l
(20
01)
(14)
Bu
sch
inge
r
et a
l
(19
94)
(15)
Bu
stos
amp C
her
ix (
1998
)(1
6) C
arli
n R
eeve
amp C
over
(19
93)
(17)
Cer
da amp
Ret
ana
(199
2) (
18)
Cer
da
et a
l
(19
94)
(19)
Cer
dan
amp P
rovo
st (
1990
) (2
0) C
eust
ers
(197
9) (
21)
Ch
agn
e B
eugn
onamp
Dej
ean
(20
00)
(22)
Ch
erix
(19
86)
(23)
Ch
erix
(19
87)
(24)
Ch
erix
amp M
adda
len
a-F
elle
r (1
987)
(25
) C
olli
ngw
ood
(198
7) (
26)
Con
way
(19
96)
(27)
Con
way
(19
97)
(28)
Cor
n (
1980
) (2
9) C
osen
s amp
Tou
ssai
nt
(198
5)
(30)
Cro
zier
P
amil
o amp
Cro
zier
(19
84)
(31)
Cu
rtis
(19
85)
(32)
Cu
shm
an
Ras
hbr
ook
amp B
eatt
ie (
1994
) (3
3)C
zech
owsk
i (1
990)
(3
4) C
zech
owsk
i (1
999)
(3
5) C
zech
owsk
i amp
Rot
kiew
icz
(199
4)
(36)
Cze
chow
ski
amp Y
amau
chi
(199
7)
(37)
Dah
bi (
1997
) (3
8) D
ahbi
amp L
enoi
r(1
998a
) (3
9) D
avid
amp W
ood
(198
0) (
40)
Dav
idso
n (
1988
) (4
1) D
avid
son
(19
97)
(42)
Dea
n (
1989
) (4
3) D
ean
amp Y
eato
n (
1993
) (4
4) D
ebou
t
et a
l
(20
03)
(45)
DeH
eer
Bac
kus
amp H
erbe
rs (
2001
) (4
6) D
ejea
n amp
Feacuten
eacuteron
(19
93)
(47)
Dej
ean
amp L
ach
aud
(199
4)
(48)
Dej
ean
et a
l
(1
993)
(4
9) D
ejea
n
et a
l
(1
994)
(5
0) D
ejea
n
Dji
eto-
Lor
don
amp D
ura
nd
(199
7) (
51)
Dej
ean
et a
l
(20
00)
(52)
Del
abie
Ben
ton
amp d
e M
edei
ros
(199
1) (
53)
Del
age-
Dar
chen
(19
74)
(54)
Del
Rio
Pes
ado
amp A
llow
ay (
1983
)(5
5) D
etra
in (
1990
) (5
6) E
lmes
(19
87)
(57)
Fed
erle
M
asch
wit
z amp
Fia
la (
1998
) (5
8) F
ern
ande
z-E
scu
dero
et a
l
(2
001)
(5
9) F
letc
her
et a
l
(1
980)
(6
0) F
oitz
ik amp
Her
bers
(20
01)
(61)
Fra
nco
eur
amp P
eacutepin
(19
78)
(62)
Gad
au
et a
l
(19
98)
(63)
Ger
st (
2001
) (6
4) G
iber
nau
amp D
ejea
n (
2001
) (6
5) G
reen
slad
e amp
Hal
lida
y (1
983)
(66
)H
arkn
ess
amp I
sham
(19
88)
(67)
Has
egaw
a (1
992)
(6
8) H
ein
ze
et a
l
(1
996)
(6
9) H
elm
s (1
999)
(7
0) H
elm
s
et a
l
(2
000)
(7
1) H
erbe
rs (
1986
) (7
2) H
erbe
rs (
1987
)(7
3) H
erbe
rs (
1989
) (7
4) p
ers
obs
ev
cite
d in
Her
bers
(19
89)
(75)
un
publ
da
ta c
ited
in
Her
bers
(19
89)
(76)
Her
bers
(19
91)
(77)
Her
bers
amp G
riec
o (1
994)
(7
8)H
erbe
rs amp
Tu
cker
(19
86)
(79)
Hof
fman
n (
1998
) (8
0) H
oumllld
oble
r (1
983)
(81
) H
oumllld
oble
r (1
984)
(82
) H
oumllld
oble
r amp
Lu
msd
en (
1980
) (8
3) H
oumllld
oble
r amp
Moumlg
lich
(19
80)
(84)
Houmll
ldob
ler
amp W
ilso
n (
1977
) (8
5) H
oumllld
oble
r amp
Wil
son
(19
86)
(86)
Houmll
ldob
ler
amp W
ilso
n (
1990
) (8
7) H
olw
ay amp
Cas
e (2
000)
(88
) Ic
hin
ose
(198
7) (
89)
Ito
Hig
ash
iamp
Mae
ta (
1988
) (9
0) J
anze
n (
1973
) (9
1) J
ayas
uri
ya amp
Tra
nie
llo
(198
5) (
92)
Joh
nso
n amp
Cro
zier
(19
98)
(93)
Kan
now
ski
(195
9) (
94)
Kas
pari
amp V
argo
(19
95)
(95)
Kel
ler
(199
1)
(96)
Kel
ler
(199
3)
(97)
var
iou
s re
fere
nce
s in
Kel
ler
(199
8)
(98)
Kel
ler
amp P
asse
ra (
1990
) (9
9) K
enn
e (1
999)
(1
00)
Kle
in (
1987
) (1
01)
Klo
tz
et a
l
(1
996)
(1
02)
Le
Mas
ne
(199
4)
(103
) L
esto
n (
1978
) (1
04)
Leacutev
ieu
x amp
Dio
man
de (
1978
) (1
05)
Lev
ieu
x (1
983)
(1
06)
Lev
ings
amp T
ran
iell
o (1
981)
(1
07)
Lie
fke
et a
l
(1
998)
(10
8) L
ongi
no
(199
1) (
109)
Lon
gin
o (2
000)
(11
0) M
abel
is (
1994
) (1
11)
Mae
der
amp C
her
ix (
2001
) (1
12)
Maj
er (
1976
) (1
13)
Mas
chw
itz
amp M
oog
(200
0) (
114)
McG
lyn
n (
1999
) (1
15)
McI
ver
(199
1) (
116)
McI
ver
amp S
teen
(19
94)
(117
) M
cKey
D (
1984
) (1
18)
Mer
cier
amp D
ejea
n (
1996
) (1
19)
Mer
cier
Len
oir
amp D
ejea
n (
1994
)(1
20)
Mer
cier
et a
l
(19
96)
(121
) M
orai
s (1
994)
(12
2) N
icke
rson
et a
l
(19
75)
(123
) O
rsquoNei
l (19
88)
(124
) O
rive
l (20
00)
(125
) P
amil
o (1
991)
(12
6) P
amil
o amp
Ros
engr
en(1
983)
(12
7) P
amil
o C
rozi
er amp
Fra
ser
(198
5) (
128)
Par
trid
ge P
artr
idge
amp F
ran
ks (
1997
) (1
29)
Pas
sera
(19
93)
(130
) P
asse
ra (
1994
) (1
31)
Pas
sera
Gil
bert
amp A
ron
(200
1)
(132
) P
eder
sen
amp B
oom
sma
(199
9)
(133
) P
eete
rs (
1993
) (1
34)
Pee
ters
amp C
rew
e (1
986)
(1
35)
Pen
g C
hri
stia
n amp
Gib
b (1
998)
(1
36)
Per
fect
o (1
994)
(1
37)
Pfe
iffe
r amp
Lin
sen
mai
r (1
998)
(13
8) P
feif
fer
amp L
inse
nm
air
(200
0) (
139)
Pfe
iffe
r amp
Lin
sen
mai
r (2
001)
(14
0) P
irk
et a
l
(20
01)
(141
) P
isar
ski
amp C
zech
owsk
i (1
990)
(1
42)
Roi
sin
et a
l
(19
86)
(143
) R
osen
gren
et a
l
(19
85)
(144
) R
owe
amp B
rist
ow (
1999
) (1
45)
Ruuml
ppel
amp H
ein
ze (
1999
) (1
46)
San
ders
amp G
ordo
n (
2000
) (1
47)
Sas
aki
Sat
oh amp
Oba
ra (
1996
) (1
48)
Sav
olai
nen
amp V
epsauml
laumlin
en (
1988
) (1
49)
Sav
olai
nen
Vep
saumllauml
inen
amp D
esli
ppe
(199
6) (
150)
Sch
mid
-Hem
pel
(198
7) (
151)
Sei
fert
(20
00)
(152
) S
mit
h-G
lase
r (1
994)
(15
3) S
nyd
er amp
Her
bers
(19
91)
(154
) S
tuar
t (1
985)
(15
5) S
tuar
t (1
987)
(15
6) S
tuar
t (1
991)
(15
7) S
un
dstr
oumlm (
1989
) (1
58)
Su
nds
troumlm
(199
3a)
(159
) T
orro
ssia
n (
1960
) (1
60)
Tor
ossi
an (
1974
) (1
61)
Tra
nie
llo
(198
2) (
162)
Tra
nie
llo
(198
9) (
163)
Tra
nie
llo
amp L
evin
gs (
1986
) (1
64)
Tsu
ji (
1988
) (1
65)
Tsu
ji amp
Yam
auch
i (1
994)
(16
6) T
suji
et a
l
(19
91)
(167
) V
asco
nce
los
amp D
avid
son
(20
00)
(168
) V
epsauml
laumlin
en
et a
l
(20
00)
(169
) W
agn
er (
1997
) (1
70)
Wag
ner
(20
00)
(171
) W
alke
r amp
Sta
mps
(19
86)
(172
) W
alof
f amp
Bla
ckit
h (
1962
) (1
73)
Way
(19
54)
(174
) Ya
mau
chi
et a
l
(19
96)
(175
) Ya
mau
chi
et a
l
(20
01)
(176
) Z
akh
arov
(19
94)
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

324
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Oec
ophy
llini
Oec
ophy
lla lo
ngin
oda
RT
HT
ER
F 82
173
A41
82
173
C41
D41
49
O49
F 41
154
n41
Oec
ophy
llini
Oec
ophy
lla s
mar
agdi
naA
US
(A
ustr
alia
) 80 1
35E
RR
FA
41 8
0 1
35C
41 8
0D
80O
41 1
35n
41 8
0
Pla
giol
epid
ini
Ano
plol
epis
long
ipes
WW
130
PA
NA
HT
129
N12
9U
129
129
n y
129
Pla
giol
epid
ini
Pla
giol
epis
pyg
mea
PAL
(Fr
ance
) 131
TE
OA
T13
1N
131
VS
131
F13
1
Pre
nole
pidi
niP
arat
rech
ina
bour
boni
caH
AN
AP
WW
T12
9N
129
U12
912
9n
129
Pre
nole
pidi
niP
arat
rech
ina
flav
ipes
HO
L (
Eas
t Asi
a [n
ativ
e] U
SA
(int
rodu
ced)
88P
AN
AH
T88
Pre
nole
pidi
niP
arat
rech
ina
long
icor
nis
WW
130
PA
NA
HT
129
N12
9U
129
O (
unic
ol)
O (
unic
ol)
O (
unic
ol)
129
n12
9
TE
RM
ITID
AE
(Is
opte
ra)
Nas
utit
erm
itina
eN
asut
iterm
es c
orni
ger
NE
O
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es n
igri
ceps
NE
O (
Pana
ma)
(Pan
ama)
4T
RR
FA
4C
4D
4F
4n
4
Nas
utit
erm
itina
eN
asut
iterm
es p
rinc
eps
AU
S (
New
Gui
nea)
142
TR
RF
A14
2C
142
F
142
n14
2
Ret
icul
iter
mit
inae
Ret
icul
iterm
es f
lavi
pes
NE
A (
USA
) 13
R
FA
13C
142
F
13n
13
Tribe
Species
Distrib
ution
Climat
icre
gion
Habita
t
Nest sit
e
Nest ty
pe
Ecolog
ical s
tatu
s
Type o
f poly
domy
Seaso
nal
Polydom
y
41
7 86
94
41 8
0
12
9
6 13
0
131
2
131
129
88
12
9
4 13
0
M P P P P P14
2
M M
Associa
ted
gyny
Colony
size
a
Syn
onym
of
P a
nti
llan
a
b
syn
onym
of
Oph
talm
opon
e
c
syn
onym
of
Har
pago
xen
us
d
syn
onym
of
M
sem
iru
fus
e
syn
onym
of
Mac
rom
isch
oid
es a
cule
atu
s
f
syn
onym
of
Hyp
ocli
nea
g
syn
onym
of
Con
omyr
ma
h
now
com
bin
ed a
s
An
onyc
hom
yrm
a n
itid
icep
s
i
syn
onym
of
Col
obop
sis
A
ll r
efer
ence
s li
sted
are
in
corp
orat
ed i
n t
he
bibl
iogr
aph
y of
th
e ar
ticl
e (
1) A
ckon
or (
1981
) 98
3)
(2)
Aco
sta
Lop
ez amp
Ser
ran
o (1
995)
(3
) A
dam
s (1
990)
99
4)
(4)
Ada
ms
amp L
evin
gs (1
987)
(5)
All
oway
et a
l
(19
82)
(6) B
ansc
hba
ch
et a
l
(19
97)
(7) B
enzi
ng
(199
1) (
8) B
eye
et a
l
(19
97)
(9) B
hat
kar
amp V
inso
n (1
987)
(10
) Boo
msm
a
et a
l
(19
90)
(11)
Bra
un
Pee
ters
amp H
oumllld
oble
r (1
994)
(12
) B
rist
ow
et a
l
(19
92)
(13)
Bu
lmer
et a
l
(20
01)
(14)
Bu
sch
inge
r
et a
l
(19
94)
(15)
Bu
stos
amp C
her
ix (
1998
)(1
6) C
arli
n R
eeve
amp C
over
(19
93)
(17)
Cer
da amp
Ret
ana
(199
2) (
18)
Cer
da
et a
l
(19
94)
(19)
Cer
dan
amp P
rovo
st (
1990
) (2
0) C
eust
ers
(197
9) (
21)
Ch
agn
e B
eugn
onamp
Dej
ean
(20
00)
(22)
Ch
erix
(19
86)
(23)
Ch
erix
(19
87)
(24)
Ch
erix
amp M
adda
len
a-F
elle
r (1
987)
(25
) C
olli
ngw
ood
(198
7) (
26)
Con
way
(19
96)
(27)
Con
way
(19
97)
(28)
Cor
n (
1980
) (2
9) C
osen
s amp
Tou
ssai
nt
(198
5)
(30)
Cro
zier
P
amil
o amp
Cro
zier
(19
84)
(31)
Cu
rtis
(19
85)
(32)
Cu
shm
an
Ras
hbr
ook
amp B
eatt
ie (
1994
) (3
3)C
zech
owsk
i (1
990)
(3
4) C
zech
owsk
i (1
999)
(3
5) C
zech
owsk
i amp
Rot
kiew
icz
(199
4)
(36)
Cze
chow
ski
amp Y
amau
chi
(199
7)
(37)
Dah
bi (
1997
) (3
8) D
ahbi
amp L
enoi
r(1
998a
) (3
9) D
avid
amp W
ood
(198
0) (
40)
Dav
idso
n (
1988
) (4
1) D
avid
son
(19
97)
(42)
Dea
n (
1989
) (4
3) D
ean
amp Y
eato
n (
1993
) (4
4) D
ebou
t
et a
l
(20
03)
(45)
DeH
eer
Bac
kus
amp H
erbe
rs (
2001
) (4
6) D
ejea
n amp
Feacuten
eacuteron
(19
93)
(47)
Dej
ean
amp L
ach
aud
(199
4)
(48)
Dej
ean
et a
l
(1
993)
(4
9) D
ejea
n
et a
l
(1
994)
(5
0) D
ejea
n
Dji
eto-
Lor
don
amp D
ura
nd
(199
7) (
51)
Dej
ean
et a
l
(20
00)
(52)
Del
abie
Ben
ton
amp d
e M
edei
ros
(199
1) (
53)
Del
age-
Dar
chen
(19
74)
(54)
Del
Rio
Pes
ado
amp A
llow
ay (
1983
)(5
5) D
etra
in (
1990
) (5
6) E
lmes
(19
87)
(57)
Fed
erle
M
asch
wit
z amp
Fia
la (
1998
) (5
8) F
ern
ande
z-E
scu
dero
et a
l
(2
001)
(5
9) F
letc
her
et a
l
(1
980)
(6
0) F
oitz
ik amp
Her
bers
(20
01)
(61)
Fra
nco
eur
amp P
eacutepin
(19
78)
(62)
Gad
au
et a
l
(19
98)
(63)
Ger
st (
2001
) (6
4) G
iber
nau
amp D
ejea
n (
2001
) (6
5) G
reen
slad
e amp
Hal
lida
y (1
983)
(66
)H
arkn
ess
amp I
sham
(19
88)
(67)
Has
egaw
a (1
992)
(6
8) H
ein
ze
et a
l
(1
996)
(6
9) H
elm
s (1
999)
(7
0) H
elm
s
et a
l
(2
000)
(7
1) H
erbe
rs (
1986
) (7
2) H
erbe
rs (
1987
)(7
3) H
erbe
rs (
1989
) (7
4) p
ers
obs
ev
cite
d in
Her
bers
(19
89)
(75)
un
publ
da
ta c
ited
in
Her
bers
(19
89)
(76)
Her
bers
(19
91)
(77)
Her
bers
amp G
riec
o (1
994)
(7
8)H
erbe
rs amp
Tu
cker
(19
86)
(79)
Hof
fman
n (
1998
) (8
0) H
oumllld
oble
r (1
983)
(81
) H
oumllld
oble
r (1
984)
(82
) H
oumllld
oble
r amp
Lu
msd
en (
1980
) (8
3) H
oumllld
oble
r amp
Moumlg
lich
(19
80)
(84)
Houmll
ldob
ler
amp W
ilso
n (
1977
) (8
5) H
oumllld
oble
r amp
Wil
son
(19
86)
(86)
Houmll
ldob
ler
amp W
ilso
n (
1990
) (8
7) H
olw
ay amp
Cas
e (2
000)
(88
) Ic
hin
ose
(198
7) (
89)
Ito
Hig
ash
iamp
Mae
ta (
1988
) (9
0) J
anze
n (
1973
) (9
1) J
ayas
uri
ya amp
Tra
nie
llo
(198
5) (
92)
Joh
nso
n amp
Cro
zier
(19
98)
(93)
Kan
now
ski
(195
9) (
94)
Kas
pari
amp V
argo
(19
95)
(95)
Kel
ler
(199
1)
(96)
Kel
ler
(199
3)
(97)
var
iou
s re
fere
nce
s in
Kel
ler
(199
8)
(98)
Kel
ler
amp P
asse
ra (
1990
) (9
9) K
enn
e (1
999)
(1
00)
Kle
in (
1987
) (1
01)
Klo
tz
et a
l
(1
996)
(1
02)
Le
Mas
ne
(199
4)
(103
) L
esto
n (
1978
) (1
04)
Leacutev
ieu
x amp
Dio
man
de (
1978
) (1
05)
Lev
ieu
x (1
983)
(1
06)
Lev
ings
amp T
ran
iell
o (1
981)
(1
07)
Lie
fke
et a
l
(1
998)
(10
8) L
ongi
no
(199
1) (
109)
Lon
gin
o (2
000)
(11
0) M
abel
is (
1994
) (1
11)
Mae
der
amp C
her
ix (
2001
) (1
12)
Maj
er (
1976
) (1
13)
Mas
chw
itz
amp M
oog
(200
0) (
114)
McG
lyn
n (
1999
) (1
15)
McI
ver
(199
1) (
116)
McI
ver
amp S
teen
(19
94)
(117
) M
cKey
D (
1984
) (1
18)
Mer
cier
amp D
ejea
n (
1996
) (1
19)
Mer
cier
Len
oir
amp D
ejea
n (
1994
)(1
20)
Mer
cier
et a
l
(19
96)
(121
) M
orai
s (1
994)
(12
2) N
icke
rson
et a
l
(19
75)
(123
) O
rsquoNei
l (19
88)
(124
) O
rive
l (20
00)
(125
) P
amil
o (1
991)
(12
6) P
amil
o amp
Ros
engr
en(1
983)
(12
7) P
amil
o C
rozi
er amp
Fra
ser
(198
5) (
128)
Par
trid
ge P
artr
idge
amp F
ran
ks (
1997
) (1
29)
Pas
sera
(19
93)
(130
) P
asse
ra (
1994
) (1
31)
Pas
sera
Gil
bert
amp A
ron
(200
1)
(132
) P
eder
sen
amp B
oom
sma
(199
9)
(133
) P
eete
rs (
1993
) (1
34)
Pee
ters
amp C
rew
e (1
986)
(1
35)
Pen
g C
hri
stia
n amp
Gib
b (1
998)
(1
36)
Per
fect
o (1
994)
(1
37)
Pfe
iffe
r amp
Lin
sen
mai
r (1
998)
(13
8) P
feif
fer
amp L
inse
nm
air
(200
0) (
139)
Pfe
iffe
r amp
Lin
sen
mai
r (2
001)
(14
0) P
irk
et a
l
(20
01)
(141
) P
isar
ski
amp C
zech
owsk
i (1
990)
(1
42)
Roi
sin
et a
l
(19
86)
(143
) R
osen
gren
et a
l
(19
85)
(144
) R
owe
amp B
rist
ow (
1999
) (1
45)
Ruuml
ppel
amp H
ein
ze (
1999
) (1
46)
San
ders
amp G
ordo
n (
2000
) (1
47)
Sas
aki
Sat
oh amp
Oba
ra (
1996
) (1
48)
Sav
olai
nen
amp V
epsauml
laumlin
en (
1988
) (1
49)
Sav
olai
nen
Vep
saumllauml
inen
amp D
esli
ppe
(199
6) (
150)
Sch
mid
-Hem
pel
(198
7) (
151)
Sei
fert
(20
00)
(152
) S
mit
h-G
lase
r (1
994)
(15
3) S
nyd
er amp
Her
bers
(19
91)
(154
) S
tuar
t (1
985)
(15
5) S
tuar
t (1
987)
(15
6) S
tuar
t (1
991)
(15
7) S
un
dstr
oumlm (
1989
) (1
58)
Su
nds
troumlm
(199
3a)
(159
) T
orro
ssia
n (
1960
) (1
60)
Tor
ossi
an (
1974
) (1
61)
Tra
nie
llo
(198
2) (
162)
Tra
nie
llo
(198
9) (
163)
Tra
nie
llo
amp L
evin
gs (
1986
) (1
64)
Tsu
ji (
1988
) (1
65)
Tsu
ji amp
Yam
auch
i (1
994)
(16
6) T
suji
et a
l
(19
91)
(167
) V
asco
nce
los
amp D
avid
son
(20
00)
(168
) V
epsauml
laumlin
en
et a
l
(20
00)
(169
) W
agn
er (
1997
) (1
70)
Wag
ner
(20
00)
(171
) W
alke
r amp
Sta
mps
(19
86)
(172
) W
alof
f amp
Bla
ckit
h (
1962
) (1
73)
Way
(19
54)
(174
) Ya
mau
chi
et a
l
(19
96)
(175
) Ya
mau
chi
et a
l
(20
01)
(176
) Z
akh
arov
(19
94)
Tab
le 1
Con
tin
ued
POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS
325
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
Thus a polydomous colony can contain queenless andqueenright nests When colonies are monogynous onlyone nest is queenright Although monogynous andpolygynous colonies may at times share common fea-tures it is necessary to clearly distinguish them in dis-cussing polydomy because polygyny and polydomymay both greatly influence social evolution in ants Inthe past because emphasis was especially placed onpolygyny the effects of polydomy have often beenunderestimated
Comparison among ant species suggests that thecomplexification of the nest structure varies continu-ously at least among terricolous ants For a largenumber of species generally characterized by smallcolony size the nest corresponds to a group of cham-bers generally vertically organized and intercon-nected by galleries (Houmllldobler amp Wilson 1990) Formost species the number of chambers and the com-plexity of the gallery network generally increase asthe colony grows In certain monodomous speciessuch as
Pachycondyla tarsata
(Dejean Beugnon ampLachaud 1993) or
Pachycondyla senaarensis
(Dejeanamp Lachaud 1994) the group of chambers of the col-onyrsquos single nest is connected by several more or lessvertically organized galleries However in the case ofboth species this gallery network is prolonged by sev-eral lateral and horizontal galleries situated severalcentimetres below the soil surface Occasionally 10 mlong or longer lateral galleries have several exitsaround each of which the workers forage over a smallarea the captured prey are transported within theselateral galleries minimizing the risk associated withexternal foraging (Dejean
et al
1993 Dejean ampLachaud 1994) Colonies of another species
Myrmi-caria opaciventris
(Kenne amp Dejean 1999) also pos-sess a group of nest chambers connected by verticalgalleries as well as lateral and horizontal galleriesbut these latter kinds of galleries lead to other groupsof nests containing brood workers and several queensthereby forming a huge polydomous nest The hightraffic of workers allowed them to transform theirsuperficial trails into trenches which are steadilytransformed over a mean of 90 days into tunnels (orlateral galleries) by adding earth little by little in theupper parts of the trenches (Kenne amp Dejean 1999)As shown by these examples the progressive passagebetween monodomy and polydomy might in certaincases originate via the exploitation of a foraging areawhich is steadily transformed into an additional nestsite certainly enabling reduction of the cost of foodtransport and of the risk of predation
Finally several types of satellite structures do notqualify as nests according to our definition The pres-ence of such structures containing workers and builtaway from the nest has also led to confusion about thedistinction between mono- and polydomy Anderson amp
McShea (2001) reviewed the various lsquoadaptive struc-tures that ants build away from the nestrsquo Theseinclude various forms of galleries outstations andconstructions dedicated to the protection of tropho-bionts nectaries or concealed food Some polydomousspecies also build certain satellite structures Thesestructures confer several adaptive benefits to the col-ony mainly in terms of defence and food foragingHowever if they contain no brood they should not beconsidered as nest sites
To summarize polydomy refers to a purely spatialand genetic proximity between several nests whereaspolycaly may be kept where convenient to refer toobserved non-aggressive behavioural interactionsbetween nests of a polydomous colony All other func-tional specializations (Anderson amp McShea 2001)including repeated architectural motifs in the utiliza-tion of space for building an ant nest may be termedas lsquomodular nestingrsquo (see section at the end of this arti-cle) There are connections between the phenomenacovered by these two concepts which may in thefuture require an all-embracing concept but this isnot the subject of the present review
R
EMINDER
OF
CONFUSING
TERMS
Anyone confronting the literature on polydomy isquickly struck by the diversity and confusion of termswhich constitutes an unavoidable source of errorsEarly work on
Formica
species illustrates this Twoterms lsquopolydomyrsquo and lsquopolycalyrsquo were frequentlyemployed either interchangeably in the same text assynonyms in translation from one language toanother or given different meanings by differentauthors Forel (1874) first introduced the term poly-caly which he defined as an arrangement of an ant col-ony in several nests each containing a more or lessindependent population necessarily constituted bybrood workers and one or several reproductivefemale(s) In the same paper Forel termed other colo-nies lacking either brood or queen in one of the nestsas polydomous After the work by Forel (1874) subse-quent studies on
Formica
introduced other terms clanfamily column family pleiad family polysectionalfamily (or anthill) cluster nest ephemeral colonysupercolony (con)federation formicarium Riesenkolo-nie community complex maternal and daughter fam-ily (or anthill or nest) multidomous colony societywith multiple nests suprafamily or conglomeration(Wellenstein 1929 Majer 1976 Houmllldobler amp Wilson1977 Leacutevieux 1983 Czechowski amp Yamauchi 1994Zakharov 1994) The term lsquocolumn familyrsquo originatedwith the theory by Zakharov (1994) stating that theintranidal population of a wood ant mound nest isdivided into separate sections like an orange Thisauthor even refers to two parallel terminologies to
326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

326
G DEBOUT
ET AL
copy 2007 The Linnean Society of London
Biological Journal of the Linnean Society
2007
90
319ndash348
describe ant social structure as the colony grows thelsquoEnglishrsquo terminology in which there is gradationfrom colony rarr polycalic colony rarr supercolony and thelsquoRussianrsquo terminology which builds a series fromfamily rarr polysectional nest rarr polycalic colony rarr truecolony rarr primary federation rarr secondary federationrarr confederation These terms have not been widelyadopted
Moreover the different units composing a polydo-mous or a polycalic colony are known by a variety ofnames adjacent nests auxiliary nests (or anthills)breeding nests bud nests colonies colony subunitsfeeding nests intercommunicating nests nestingunits satellite nests shelters secondary nests sistercolonies subnests and subsidiary nests (Weber 1935Waloff amp Blackith 1962 Majer 1976 David amp Wood1980 Leacutevieux 1983 Jayasuriya amp Traniello 1985Traniello 1989 McIver amp Steen 1994 Morais 1994Zakharov 1994 Klotz Reid amp Klotz 1996 Bansch-bach Levit amp Herbers 1997 Billick 1999 Dejeanet al 2000 Fernandez-Escudero Seppauml amp Pamilo2001) These terms generally used to describe nests ofpolydomous colonies have also been used to refer tovarious kinds of lsquooutstationsrsquo
CONFUSING SITUATIONS
A second source of confusion about polydomy is thefact that the degree to which it is expressed oftendepends on factors such as colony size (Majer 1976Tsuji 1988) Many terms were used by early research-ers to describe or distinguish categories in a contin-uum of social structure The large range of colony sizeencountered in some Formica species is responsible forthe profusion of terms used to describe the differentpopulation levels and social structures observed(Zakharov 1994 see also above) Some species havebeen described as polydomous only for a small propor-tion of all colonies (eg 23ndash137 of all colonies inPristomyrmex pungens Tsuji 1988) whereas in otherspecies such as Oecophylla longinoda polydomyappears to be constant Several species of Formica alsoexhibit two kinds of colony structure Some coloniesare monodomous and monogynous whereas others arepolydomous and polygynous (Pamilo 1991) Howeversuch a clear relationship between queen number andsocial structure is far from being a generality amongants and numerous monogynous species are also poly-domous (see below)
Two other particular situations linked to specificbiological traits could be confused with true polydomyFirst mixed colonies could introduce some confusionbecause they may involve two polydomous species(eg Formica sanguinea and Formica cinerea cinereaCzechowski amp Rotkiewicz 1997) a monodomous and apolydomous species or two monodomous species (ie
Orivel Errard amp Dejean 1997) Similar consider-ations apply to socially parasitic species and theirhosts (Del Rio Pesado amp Alloway 1983) Second somespecies of the ponerine genus Centromyrmex whichare specialized termite predators inhabit the termi-taries they exploit occupying lodges that may be spa-tially separated (Delabie 1995 Dejean amp Feacuteneacuteron1999) Such nesting habits called termitolesty maythus lead to lsquoa certain kind of polydomyrsquo (Dejean ampFeacuteneacuteron 1999) but are not very well known
POLYDOMY AS A BY-PRODUCT OF POLYGYNY
Polygyny may be the predominant social structure insocial insects especially among ants (Keller 1993)and interspecific comparisons suggest that polydomyin polygynous species is often a secondary evolutionarystep that accompanied or followed evolution of poly-gyny from a monogynous ancestor However althoughpolydomy is often associated with polygyny (Holldoumlbleramp Wilson 1977 Rosengren amp Pamilo 1983 Ross ampFletcher 1985 Keller 1991) many monogynous butpolydomous species are known (Way 1954 Traniello ampLevings 1986 Ichinose 1987 Snyder amp Herbers1991 Buschinger Klein amp Maschwitz 1994 CerdaDahbi amp de Haro 1994 for additional examples seeTable 1) As noted by Houmllldobler amp Wilson (1977) lsquothecorrelation (between polygyny and polydomy) is veryweakrsquo It appears that lsquomany monogynous ant speciesare also polydomous while a few polygynous ones aremonodomousrsquo (Houmllldobler amp Wilson 1977)
The frequent association of polygyny and polydomymay mean that similar factors favour both traits Thepredominant environmental influence determiningthe degree of polygyny is the cost of dispersal (Keller1995) A high cost of dispersal could also promote poly-domy In polygynous populations polydomy is the con-sequence of lsquobuddingrsquo a mechanism of local dispersaleffected by groups of workers accompanied by one ormore queens The probability of success is usuallyhigher (lower cost of dispersal) than when foundressesdisperse alone to attempt to establish independent col-onies In such cases polydomy is associated withpolygyny either facultative (Higashi 1979) or obligate(eg in Formica polyctena Rosengren amp Pamilo 1983)Here polydomy reduces conflicts among maturequeens of the same polygynous nest In some casesbudding followed by separation has replaced indepen-dent establishment by foundresses as the prevalentprocess of founding new colonies (a phenomenoncalled lsquofissionrsquo) In these cases a polydomous colonystructure may simply be a transient stage (eg inF polyctena Rosengren amp Pamilo 1983)
Finally acquisition of polydomy could be seen as abet-hedging strategy if one nest is destroyed others
POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 327
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
remain available to the colony This advantage appliesalso to monogynous colonies (if the queen survivesdestruction of the queenright nest she could moveinto one of the other nests) but is likely to be of great-est importance in polygynous colonies If one queen-right nest is destroyed related queens remain alive inother parts of the colony However in Formica trun-corum even when they include tens of flourishingnests and hundreds of queens polydomous and polyg-ynous colonies may succumb within a few years andthus appear unexpectedly vulnerable (R Rosengrenpers comm) In this case ecological (increased rate oftransmission of parasites due to very high density ofnests) or genetic factors (selfish behaviours caused bythe relatively low relatedness between queens) may beresponsible We are aware of no dataset suggestingthat polydomous structure in polygynous speciesincreases the probability of survival of queens andorcolonies
THE SYNDROME OF POLYDOMY IN ANTS
Features commonly associated with polydomy includethe overdensity of same-species neighbours the exist-
ence of internest distances smaller than thoseobserved in monodomous populations and the overallpattern of nest aggregation (Stevens 2000) Are othertraits particularly frequent in polydomous ants Weattempted an exhaustive review of the literature tolist all ant species which by our criteria can bedescribed as exhibiting facultative or obligatorypolydomy Bibliographic research was facilitatedby the ant literature database FORMIS (httpcmaveusdaufledu~formis) Current Contents andseveral personal literature databases kindly madeavailable by individuals (see Acknowledgements) Weexamined whether various life-history traits were sys-tematically associated with the presence of polydo-mous structure and whether they covaried with thetype of polydomy taking into account only the traitsfor which data were available for more than 50 of allcensused species (Table 2) The results of this revieware presented in Table 1
Polydomy has been recorded at least once for a totalof 166 ant species belonging to 49 genera Weattempted to determine whether ecological or otherbiological traits were associated with polydomy toform some recognizable syndrome Our objective was
Table 2 List of the traits examined
Trait Abbreviation Meaning Trait Symbol Meaning
Distribution ETH Ethiopian Nest type C Self-constructionPAL Palearctic E ExcavationHOL Holarctic N Natural cavityNEA Nearctic Ecological status D DominantNEW New world U UnicolonialNEO Neotropical S Slave-making speciesAUS Australian N Sub- and nondominantORI Oriental (= Asian tropics) VS Variable statusAAS Australasian Type of polydomy F Facultative (size-dependen)WW Cosmopolitan O Obligatory (unicolonial)
Climaticregion
TE Temperate Seasonal polydomy N NoER Temperate + tropical Y YesST Subtropical Associated gyny M Strict monogynyTR Tropical P Strict polygynyPAN Panclimatic NL No link between number of
queens and nestsHabitat AH Associated with humans W Without queen
MZ Mediterranean forest Colony size 1 100OA Open areas (maximum number
of workers)2 500
RF Rain forest 3 1000TF Temperate and boreal
forest4 5000
Nest-sitelocation
A Arboricolous 5 10 000T Terricolous 6 100 000M Both types 7 1000 000
8 gt 106
328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

328 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
not to draw a comparison between polydomous andmonodomous species Polydomy is an evolutionarilylabile trait that has evolved numerous times indepen-dently Polydomy appears not to be linked to any nar-row set of ecological or social conditions Althoughsome traits appeared frequently associated no syn-drome emerged (Fig 1) Ant species that express poly-domous colonial structure do not always present thesame set of associated life history traits whether con-sidering the habitat range the characteristics of nestsor ecological and social traits Polydomy is present inall the main subfamilies (Ponerinae Pseudomyrmeci-nae Myrmicinae Aneuretinae Dolichoderinae andFormicinae) and on all continents The main biomesconcerned are forested habitats both tropical (385)and temperate (283) Open areas are a minorityand some habitats (eg desert swamp) were notrepresented in our sample (they are doubtless alsounderrepresented among ant studies) Half of thepolydomous species can be characterized as ecologi-cally dominant but all other positions in competitivehierarchies are also represented (subdominant subor-dinate) and it should be kept in mind that competitivehierarchies are relative rather than objective notions(the same species can be dominant or not dependingon ecological conditions) Polydomous species includeboth terricolous and arboricolous ants and nest sitescan be of various types (natural cavities self-con-structed nests or excavations) Among species inwhich polydomy has been recorded it is a facultativetrait in 837 of species (among or even within popu-lations) Seasonal polydomy is rare having been dem-onstrated only in 10 of polydomous species Finallyand most importantly polydomy is not inordinatelyfrequently associated with polygyny confirming theconclusion of Houmllldobler amp Wilson (1977) Half of thepolydomous species for which gyny status has beendescribed are apparently always monogynous
To examine whether polydomy is associated withdifferent suites of traits in different lineages theextent of differentiation among lineages (congenericspecies genera subfamilies) was assessed by perform-ing principal component analysis (PCA) using thePRINCOMP procedure in SAS version 80 (SAS Insti-tute) The analysis was conducted on a correlationmatrix obtained after transformation of the data pre-sented in Table 1 using the optimal scoring method ofFisher (1938) that assigns scores to each class (level)of the variable character or numeric (SAS InstituteInc 1999) A pattern of species differentiation levelwas revealed by PCA The first three axes explained825 of the total variance and allowed discriminationof three groups on the basis of how polydomy isexpressed species with seasonal polydomy (type Ispecies with nonseasonal polydomous structure (typeII) and unicolonial species (type III) (Fig 2) None of
the other traits considered nor the lineages couldexplain differentiation At the generic level the firstthree axes explained 776 of the total variance(Fig 3) The first axis allowed the discrimination ofgenera including unicolonial species (type C) from allother genera Again no discrimination between sub-families was evident The axis δ (a linear combinationof axes 1 and 2) allowed discrimination betweengroups mainly in relation to habitat and nest-sitelocation (ground or tree nesting) type A groupstogether arboreal genera from the New World (thesegenera are principally composed of monogynous spe-cies) and type B groups together terricolous generaprincipally from the Palearctic region and tropicalAsia However it should be noted that estimation ofthe mean point for each genus may have introducedsome bias because this is a narrow way to synthesizeall characteristics of species from one genus in onlyone set of traits
In summary similar traits were correlated withpolydomy in monogynous and in polygynous speciesWhatever the taxonomic level considered polydomywas always associated with distributional or ecologicalcriteria but never with subfamily or gyny status Inpolygynous species the potential link between poly-domy and foraging strategy or territoriality (when dis-persal success of females is low) is always hidden bythe overlap of polydomy and polygyny thus in thesecases polydomy may be parsimoniously considered asa by-product of polygyny In monogynous species poly-domy may be more clearly linked to foraging strategyandor may be a consequence of strong queenndashworkerconflict Finally there are no invariant correlates ofpolydomy Transitions between monodomy and poly-domy seem to have occurred frequently apparently inresponse to various ecological factors Polydomy is cer-tainly a very labile life history trait which may varyextremely depending on the ecology of the ant speciesand on environmental variation A summary of the dif-ferent hypotheses explaining the potential evolution-ary causes andor consequences of polydomy isproposed in Table 3 and these hypotheses are dis-cussed thereafter
ECOLOGY OF POLYDOMY
Polydomy can confer numerous ecological advantagesYet without phylogenetic information it is difficult todetermine which of these could be selective advan-tages that initially favoured polydomy and whichwere later consequences of polydomy The latter caninclude direct adaptive consequences of polydomy newadaptive traits whose evolution was favoured by theacquisition of polydomy or simply by-products of hav-ing multiple-nest structure Polydomous structureallows nests to be dispersed shortening foraging
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292
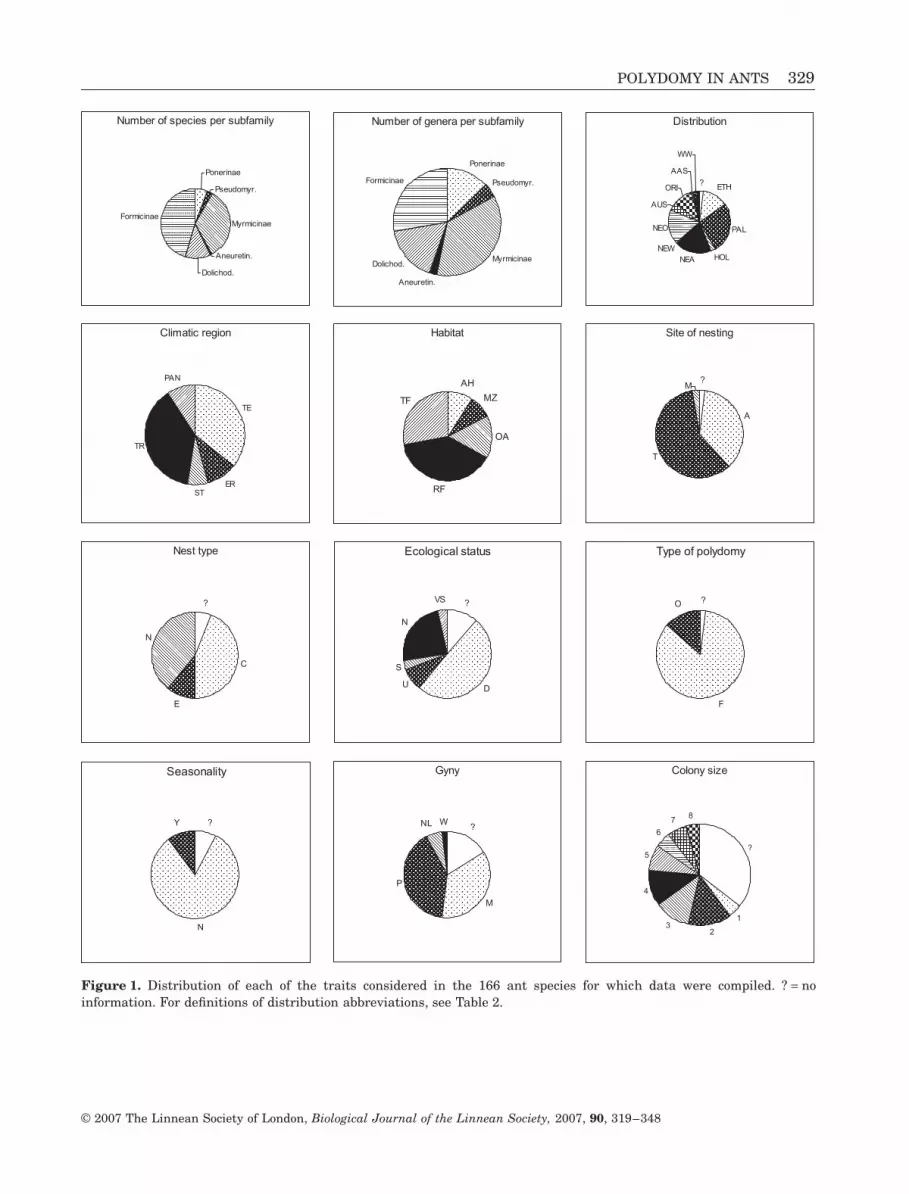
POLYDOMY IN ANTS 329
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Figure 1 Distribution of each of the traits considered in the 166 ant species for which data were compiled = noinformation For definitions of distribution abbreviations see Table 2
Distribution
ETH
PAL
HOLNEANEW
NEO
AUS
ORI
AAS
WW
Ecological status
DU
S
N
VS
Type of polydomy
F
O
Climatic region
TE
ERST
TR
PAN
Site of nesting
A
T
M
Seasonality
N
Y
Habitat
AH
MZ
OA
RF
TF
Nest type
C
E
N
Gyny
M
P
NL W
Colony size
1
23
4
5
6
7 8
Number of species per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
Number of genera per subfamily
Ponerinae
Pseudomyr
Myrmicinae
Aneuretin
Dolichod
Formicinae
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292
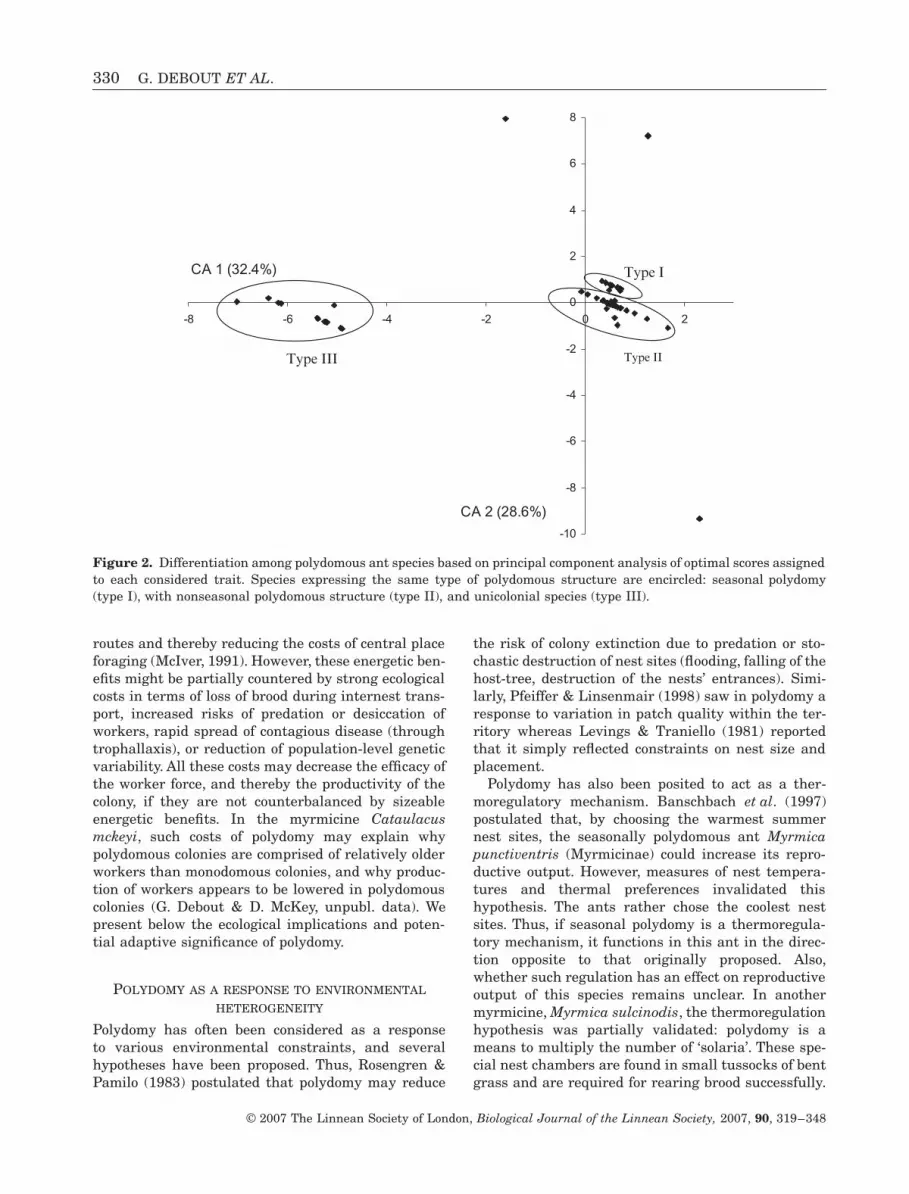
330 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
routes and thereby reducing the costs of central placeforaging (McIver 1991) However these energetic ben-efits might be partially countered by strong ecologicalcosts in terms of loss of brood during internest trans-port increased risks of predation or desiccation ofworkers rapid spread of contagious disease (throughtrophallaxis) or reduction of population-level geneticvariability All these costs may decrease the efficacy ofthe worker force and thereby the productivity of thecolony if they are not counterbalanced by sizeableenergetic benefits In the myrmicine Cataulacusmckeyi such costs of polydomy may explain whypolydomous colonies are comprised of relatively olderworkers than monodomous colonies and why produc-tion of workers appears to be lowered in polydomouscolonies (G Debout amp D McKey unpubl data) Wepresent below the ecological implications and poten-tial adaptive significance of polydomy
POLYDOMY AS A RESPONSE TO ENVIRONMENTAL HETEROGENEITY
Polydomy has often been considered as a responseto various environmental constraints and severalhypotheses have been proposed Thus Rosengren ampPamilo (1983) postulated that polydomy may reduce
the risk of colony extinction due to predation or sto-chastic destruction of nest sites (flooding falling of thehost-tree destruction of the nestsrsquo entrances) Simi-larly Pfeiffer amp Linsenmair (1998) saw in polydomy aresponse to variation in patch quality within the ter-ritory whereas Levings amp Traniello (1981) reportedthat it simply reflected constraints on nest size andplacement
Polydomy has also been posited to act as a ther-moregulatory mechanism Banschbach et al (1997)postulated that by choosing the warmest summernest sites the seasonally polydomous ant Myrmicapunctiventris (Myrmicinae) could increase its repro-ductive output However measures of nest tempera-tures and thermal preferences invalidated thishypothesis The ants rather chose the coolest nestsites Thus if seasonal polydomy is a thermoregula-tory mechanism it functions in this ant in the direc-tion opposite to that originally proposed Alsowhether such regulation has an effect on reproductiveoutput of this species remains unclear In anothermyrmicine Myrmica sulcinodis the thermoregulationhypothesis was partially validated polydomy is ameans to multiply the number of lsquosolariarsquo These spe-cial nest chambers are found in small tussocks of bentgrass and are required for rearing brood successfully
Figure 2 Differentiation among polydomous ant species based on principal component analysis of optimal scores assignedto each considered trait Species expressing the same type of polydomous structure are encircled seasonal polydomy(type I) with nonseasonal polydomous structure (type II) and unicolonial species (type III)
-10
-8
-6
-4
-2
0
2
4
6
8
-8 -6 -4 -2 0 2
CA 1 (324)
CA 2 (286)
Type II
Type I
Type III
POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 331
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Because these tussocks are scattered within a patchhaving multiple nests may increase the rearing capac-ity and thus the reproductive output of the colony(Pedersen amp Boomsma 1999) This thermoregulatorymechanism was also found in highly polydomous col-onies of F truncorum that live on open islands in theGulf of Finland (R Rosengren pers comm)
POLYDOMY AND THE EFFICIENCY OF RESOURCE CAPTURE
By acquiring a polydomous structure a colony mayincrease its rate of capture of resources (food or nestsites) by the expansion of its foraging area andincreased efficiency of foraging By allowing the colonyto forage over a greater area polydomy also allows thediversification of food resources and thereby strength-ens opportunism in foraging and thereby the stabilityof the colonyrsquos food supply Social insect colonies havefrequently been considered as central-place foragerssimilar to some solitary animals (Houmllldobler amp Lums-den 1980) However polydomous ant societies divergefrom the classic central-place model because the dif-ferent nests of a colony are often not aggregated in onecentral place They are instead decentralized through-
out much of the colonyrsquos territory (ie the area that thecolony occupies exclusively and defends againstintraspecific and often interspecific intruders Houmlll-dobler amp Lumsden 1980) Through this switch fromcentral-place to decentralized or dispersed central-place foraging behaviour the whole system becomeshighly flexible in the distribution of nests and its allo-cation of workers brood and resources among neststhroughout a colonyrsquos foraging area Such a colonyorganization should confer advantages whenresources are patchily distributed (McIver 1991Holway amp Case 2000) In several species from verydifferent subfamilies such as Lasius neoniger (For-micinae) (Traniello amp Levings 1986) M punctiventris(Banschbach amp Herbers 1996a) or Linepithemahumile (Dolichoderinae) (Holway amp Case 2000) poly-domy is proposed to have arisen as an adaptationrelated primarily to foraging ecology For Camponotusgigas (Formicinae) the link between polydomy andforaging organization has direct morphological andsocial consequences because a group of specialisttransport worker ants carries food from lsquosourcersquo nestsat the periphery to the central queenright lsquosinkrsquo nestThese transporter ants form a physical subcasteamong the minors and behave according to predictions
Figure 3 Differentiation among polydomous ant genera based on principal component analysis of optimal scores assignedto each considered trait Genera expressing the same set of ecologicaldistributional traits are encircled and the differen-tiating traits of each group are indicated arboreal genera from the New World (type A) terricolous genera principallyfrom the Palearctic zone and Asia (type B) and unicolonial species (type D)
-6
-5
-4
-3
-2
-1
0
1
2
-3 -2 -1 0 1 2 3 4 5 6
CA 1 (423)
CA 2 (198 )
Type A
Type B Type C
332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

332 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
of central-place foraging theory (Pfeiffer amp Linsen-mair 1998)
By this strategy foraging costs are minimizedthrough a reduction of travel distances and times andthereby also of exposure to natural enemies The par-titioning of the territory may reduce the loss of prey toother competitors (Traniello amp Levings 1986) as wellas the costs of prey transport from distant capturepoints to a single central nest deep within the territory(McIver 1991) Thus polydomous ants are thought toachieve energetic savings by decentralization (Houmlll-dobler amp Lumsden 1980) especially by reducing theoverlap in the individual foraging paths (Traniello ampLevings 1986 Davidson 1997) thereby increasingtheir foraging efficiency competitive ability and effec-tiveness in defending resources (Cherix amp Bourne1980 Rosengren 1986)
However competition for food cannot alone explainsome traits frequently associated with polydomy suchas the overdensity of same-species neighbours theexistence of shorter internest distances and the over-all pattern of nest aggregation (Stevens 2000) Limi-tation by a resource other than food such as theavailability of nest sites can better explain these pat-terns and polydomy might thus be a response to highor low (depending on the costs of dispersal) density of
suitable nest sites and their size distribution(Herbers 1989)
Polydomy thus appears to confer advantages inthe energetics of foraging and in the exploitation ofheterogeneous environments These advantages canexplain the high ecological success of polydomous spe-cies or societies Ecologically dominant ants are oftenpolydomous and the foraging behaviour of these spe-cies suggests that nest relocation and the mainte-nance of multiple nests both play a role in theircompetitive prowess Among well-known polydomousdominant ant species are the tropical O longinoda(Way 1954) and the temperate F truncorum (Rosen-gren 1986 Sundstroumlm 1993a) and other Formicaspecies (Cherix Werner amp Catzeflis et al 1980Rosengren amp Pamilo 1983 Savolainen amp Vepsaumllaumlinen1989 Punttila 1996)
POLYDOMY AS A CHARACTERISTIC TRAIT OF TRAMPINVASIVE ANT SPECIES
Remarkable examples of polydomous dominant antsare provided by invasive species such as Pheidolemegacephala (Myrmicinae) (Houmllldobler amp Wilson1977) Wasmannia auropunctata (Myrmicinae) (Clarket al 1982) Lasius neglectus (Formicinae) (Espadeler
Table 3 Hypotheses to account for the evolution of polydomy and their implications for social organization
Hypothesis Possible implications for social organization
Expansion of the foraging area increase of the efficiency of foraging (2)
If there is resource heterogeneity in space in time appearance of lsquosinkrsquo nests and lsquosourcersquo nests evolution of a special subcaste of transporter ants which carry the food between lsquosourcersquo and lsquosinkrsquo nests (6)
Decrease of the risk of colony extinction due to predation or stochastic destruction of nest sites (7)
Poor flow of information between nests (except for ant species with long-distance recruitment behaviour andor moderate colony size) too small worker groups unfavourable defence opportunities
Constraints on nest size and placement (4)
In polygynous species= by-product of the mode of colony reproduc-
tion (budding) andor of unicolonialityAllows the formation of new autonomous colonies by budding or fission
of the old polydomous colony (5) As the cost of producing sexuals and the cost of dispersal are decreased polydomous and polygynous species should show reduced nuptial flight activity low dispersal weak territoriality or colony reproduction by budding (9 10)
= combination of polygyny with nest site insta-bility (3 10)
Empirical evidence in Lasius reduced nuptial flight activity low dispersal weak territoriality polygyny or colony reproduction by budding cited in (8)
In monogynous species= adaptation related primarily to foraging
ecology especially when prey items are unpredictable in both space and time (8)
In queenless nests workers may escape queen control over sex allocation and achieve a sex ratio closer to their own optimum (1)
Evolution of polygyny
All references listed are incorporated in the bibliography of the article (1) Herbers (1984) (2) Holldoumlbler amp Lumsden 1980(3) Houmllldobler amp Wilson (1977) (4) Levings amp Traniello (1981) (5) Pedersen amp Boomsma (1999) (6) Pfeiffer amp Linsenmair(1998) (7) Rosengren amp Pamilo (1983) (8) Traniello amp Levings (1986) (9) Wilson (1953) (10) Yamauchi et al (1996)
POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 333
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
amp Rey 2001) the argentine ant L humile (Doli-choderinae) (Reuter et al 2001 Tsutsui amp Case 2001Giraud et al 2002) and the polygynous form of Sole-nopsis invicta (Myrmicinae) (Passera 1994) for areview of invasive unicolonial species see also Chap-man amp Bourke (2001) Unicolonial polydomy exhibitedby these ant species appears to have facilitated theirspread Holway amp Case (2000) propose the followingscenario At introduction the population size of newinvasive species is generally low leading to a bottle-neck and low genetic variation of the introduced pop-ulations as in the Argentine ant L humile introducedin North America (Tsutsui et al 2000 Tsutsui ampCase 2001) However Giraud et al (2002) proposedthat the spread of the Argentine ant in Europe did notfollow a small number of introduction events (bottle-neck) and that the lack of aggressiveness could beinstead due to a selected decrease of allelic diversity atthe recognition loci a mechanism allowing colonies ofthe introduced species to rapidly settle in the newarea Because of this low differentiation lowerintraspecific levels of aggression are generallyobserved tending to unicoloniality Here polydomy isclosely tied to the low level of genetic variation (lack ofcolony closure) and this specific trait generally bringsa high ecological advantage in terms of resource cap-ture Polydomy associated with unicoloniality hasprobably been the key social trait that led to successfulinvasion and the replacement of native ants (Holwayamp Case 2000)
CONSEQUENCES OF POLYDOMY FOR SOCIAL EVOLUTION
POLYDOMY AND WITHIN-COLONY CONFLICTS
Social insect colonies are not the completely coopera-tive units depicted by traditional descriptions(Backus 1993) Instead there exists a subtle equilib-rium between cooperation and competition among allmembers of an ant colony Each individual ant mightbehave to maximize its own inclusive fitness (classicalfitness plus kinship components) For example it hasbeen demonstrated that nepotistic behaviours canarise not only between reproductive females but alsobetween workers that are able to favour their ownclose kin when rearing eggs and larvae in coloniesheaded by several queens (Hannonen amp Sundstroumlm2003) Such selfish or nepotistic behaviour may befavoured up to the point where the resulting decreasein cooperation and social cohesion of the colony leadsto reduction of survival andor reproductive output(Keller 1995)
In a polydomous colony due to the greater orlesser independence of each nest there is a poten-tial to exacerbate conflicts among reproductive
females among workers themselves or betweenworkers and their mother queen(s) or to generatenovel outcomes of conflicts that already exist Con-flicts over traits such as allocation of reproductiveeffort to males or who produces males could beexpressed as competition between lsquosinkrsquo and lsquosourcersquonests These conflicts could arise simply because ofthe physical barrier between individuals of the dif-ferent parts of the colony Distance between nestshinders colony homogenization because it increasespredation on workers sexuals and brood andlimits chemical communication between individualsthereby affecting the control that some colony mem-bers may exert on others Because polydomy pro-vides a novel stage upon which these variousconflicts (but essentially queenndashworker conflict) areplayed out polydomous ant colonies are good mod-els to study the equilibrium between cooperationand competition by providing opportunities to exam-ine the levels at which selection acts
However polydomous ant species have developedparticular behaviours such as exchange of queens orworkers foraging cooperation or exchange of cuticu-lar compounds between nests that may function todecrease the conflicts potentially exacerbated by poly-domy and thus to create the necessary social environ-ment to harmonize individual and group interestsThese behaviours tend to diminish genetic variationamong nests thereby minimizing the potential forgenetic conflict The question remains whether thereduction of conflict is an evolved function of thesebehaviours or whether it is a convenient by-productof behaviours whose adaptive significance lieselsewhere
A NEW LEVEL OF SELECTION THE NEST-LEVEL
Any study of how insect societies are structured pre-sents a levels-of-selection problem Natural selectionultimately acts on genes whatever the level of orga-nization considered and selection at the level of indi-viduals is usually congruent with selection acting atthe level of genes because an individual is a society ofcooperating genes Selection can be said to occur at asupra-individual level if the gain in fitness for thegenes at this level is great enough to counterbalance aloss in fitness at the individual level In consequencesome authors have proposed that an ant colony couldbe likened to a lsquosuperorganismrsquo with its own colony-level fitness When we consider a polydomous struc-ture a new level of organization the nest-level mayemerge Whether selection occurs at this new level is apertinent question when studying polydomous societ-ies Indeed in some polydomous ants it has been sug-gested that lsquonest-levelrsquo allocation is subjected tostronger selection than is allocation at the colony level
334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

334 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
(Herbers 1984 Snyder amp Herbers 1991) Banschbachamp Herbers (1996b) demonstrated that resource alloca-tion to maintenance (proportion of resources allocatedto workers rather than reproduction) and sex alloca-tion (allocation of reproductive effort between thesexes) were both strongly dependent on the number ofworkers in the nest and less strongly dependent onqueen presenceabsence in the nest They suggest thatonly nest-level traits are important in determiningvariation in fitness
However not all investigators agree about thepertinence of the lsquonest-levelrsquo in analysing selectionon social traits Sex allocation for example appearsto remain subject to selection at the colony leveldespite the polydomous structure of the colony Forexample in Technomyrmex albipes (Dolichoderi-nae) whose mature colonies are highly polygynousand polydomous a colony functions as a wholeentity Between-colony variances of allocation levelsare always larger than the within-colony-internestvariances The homogeneity of life-history traitsbetween nests is probably due to the frequentmovements of workers between the nests of a col-ony (Tsuji amp Yamauchi 1994 Dahbi et al 1996)Nevertheless polydomous ant colonies representunique opportunities to explore how selection mightoperate hierarchically
MODIFICATIONS IN REPRODUCTIVE AND SEX ALLOCATIONS
Whatever the colony structure reproductive strat-egy may vary in time depending on conditions ofcompetition availability of nest sites (space satura-tion) and access to resources However a multineststructure will intrinsically lead to modifications inresource allocation and of the reproductive strate-gies adopted by the colony Whereas precise theoreti-cal models treat the effects on resource allocation oftraits such as polygyny multiple mating or workerreproduction (Crozier amp Pamilo 1996 Frank 1998)the lack of theory specifically treating polydomyhampers progress in this domain Looking at poly-domous colonies as metapopulations could help atleast for polygynous societies Indeed the poly-domous colony can be seen as the intermediate levelin a metapopulation hierarchy in which the highestlevel is constituted by the set of conspecific colonieswithin an area and the lowest level is represented bythe population of reproductive queens within a singlenest However metapopulation theory has yet to beapplied to allocation strategies in polydomous colo-nies We list below all the theoretical predictions ofwhich we are aware that have been made aboutreproductive resource allocation or sex allocation inpolydomous ant species
Theoretical predictions on the allocation of resources to reproductionAlthough most work on queenndashworker conflict hasfocused on sex allocation patterns other allocationdecisions may also be subject to conflict Pamilo (1991)predicted that queenndashworker conflict over the divisionof resources between reproduction and growth (repro-ductive allocation) may occur in social insects butuntil recently this conflict over allocation to workersvs sexuals (growth vs reproduction) had not been wellstudied nor further developed by theoreticians(Backus 1995) Contrary to the predictions of Pamilo(1991) Bourke amp Chan (1999) demonstrated that aconflict over relative resource allocation to sexuals vsworkers is not expected to occur under the conditionsof monogyny monoandry and worker sterilitybecause under these circumstances queen and work-ers can only invest in sexuals (new queens and males)derived from the colony queen At population sexratio equilibrium potential conflict over reproductiveresource allocation is absent because both partiesmaximize fitness by maximizing the colonyrsquos total out-put of these sexuals This does not prevent potentialconflicts over sex allocation from occurring Resolvingthe apparent contradiction between the results ofPamilo (1991) and Bourke amp Chan (1999) HerbersDeHeer amp Foitzik (2001) developed a model thatshows the dependence of reproductive-allocation con-flict on sex-allocation conflict The queenndashworker con-flict over reproductive allocation depends not only onthe existence of a conflict over sex allocation but alsoon the existence of variation in sex ratios across dif-ferent colonies in a population
On the other hand because the interests of queensand workers differ over when queens should bereplaced in the case of polygyny workers could biasallocation in favour of rapid reproduction and readopt-ing of daughter queens Indeed in the case of recur-ring queen turnover workers always favour queenreplacement sooner than the queen since they tradesiblings (r = 05) for offspring of a full sister (r = 0375)whereas the queen trades offspring (r = 05) for grand-offspring (r = 025) (Crozier amp Pamilo 1996 Bourke ampChan 1999) In other words exacerbated differencesin the relative allocation of resources to new queensand new workers should be observed between queen-less and queenright nests in polygynous colonies ifthe queen controls the investment in sexuals in queen-right nests These differences can also occur underworker control because variations of sex ratio areexpected between queenless and queenright nests (seebelow) and a male-biased sex ratio can acceleratequeen replacement by workers (Crozier amp Pamilo1996) Reuter amp Keller (2001) proposed a model onpotential conflicts over resource allocation in the pro-duction of workers males and gynes that considered
POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 335
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
the power that queens and workers have to manipu-late resource allocation Taking into account the selec-tion pressures acting on queens workers and larvaethey showed that queenndashworker conflict occurs oversex allocation but that the investment in workers isbelow both partiesrsquo optima The strongest conflict isbetween queens and workers on the one hand andfemale larvae on the other over the developmentalfate of the latter
The effects of polydomy on the conflict over resourceallocation between production of workers and of sex-uals have never been considered despite the fact thatthis allocation is certainly a fundamental decision inthe life history of a colony Indeed workers areassumed to be associated with colony survivorship andthey represent the interface between the colony andthe environment Conflicts may arise when queensand workers have options other than ones for whichtheir fitness optima coincide For example when someof the nests of a colony are queenless workers mayhave greater opportunity to alter significantly the allo-cation pattern because in these nests one of the twoparties is absent Only one empirical study hasdetected queenndashworker conflict over allocation togrowth vs reproduction in Leptothorax longispinosus(Backus 1993) Banschbach amp Herbers (1996b)showed strong differences between populations inworker allocation for M punctiventris but they wereunable to link these variations to any social (queennumber colonial structure) or ecological parametersHerbers et al (2001) performed new analyses of thedata sets for both species based on the predictions oftheir new model Variation in reproductive allocationas a function of sex allocation did not show clear pat-terns but the approach underlines the need for fur-ther tests of conflict theory that take into account theeffects of sex allocation and of colony size on reproduc-tive allocation
Theoretical predictions on sex allocationAlthough inclusive fitness theory predicts that partialor complete worker control of sex allocation leads tooverall female-biased sex ratios authors generallyagree that male-biased investment at the colony-levelis expected under polydomy Three hypotheses havebeen advanced
1 In polygynous societies polydomy is often associ-ated with nest-founding by budding (a queen leavesthe nest accompanied by a few workers) or colonyfission (a mature nest lsquoexplodesrsquo to form several groupsof workers each with one or more queens) The latterphenomenon is so far known exclusively from driverants (Dorylinae) In both cases dispersal range offemales is much smaller than that of males Even ifthese phenomena should mitigate sib-competition in
the natal nest they could also lead to sib-competitionbetween nest-founding females (a form of localresource competition) if the local habitat patch is sat-urated with nests or if the resource level in the hab-itat patch is low Under these conditions colonies areexpected to produce a small number of new queensand a large number of males (Clark 1978 Craig 1980Bulmer 1983)
2 In species that reproduce only through swarming(budding or colony fission) the investment in workersassociated with the nest-founding gynes should becounted as an investment in the female function whencalculating the sex investment ratios (Macevicz1979) Because colonies require only one or a fewqueens this has led to the prediction of highly male-biased numerical sex ratios amongst the alates them-selves Because the workers that participate in bud-ding cannot be entirely included in investment inproduction of females (these workers will haveinvested some portion of their lives before budding inraising males) estimating a true investment ratiobecomes rather problematic Thus sex ratio amongstalates remains the easiest although biased sex allo-cation estimator (Nonacs 1993)
3 In the same way diploid males although generallysterile are still in practice recorded as males when thesex ratio is assessed because they often cannot bedistinguished from other males In the case of queencontrol of the sex ratio these diploid males could evenbe counted as females because they are lsquointendedrsquofemales in the sense that they originated from eggsfertilized by the queen In ants heterozygosity at onespecific locus determines females haplozygosity deter-mines males and homozygosity determines diploidmales (Bourke amp Franks 1995) In polygynous poly-domous colonies inbreeding might be substantial ineach of the subnests leading to higher frequencies ofdiploid males than in monodomous colonies For exam-ple in some polydomous island populations ofF truncorum up to 10 of males were diploid (Pamiloet al 1994)
Two other considerations although not necessarilyleading to predictions of a male-biased sex ratio leadto the related prediction that sex ratios in polydomouscolonies should not be as female-biased as expectedfrom relatedness asymmetry (Trivers amp Hare 1976)
4 Polydomous societies are often polygynous As thequeenndashworker conflict is diminished then even in thecase of total worker control we should expect a sexratio closer to 1 1 than 3 1 (Trivers amp Hare 1976Bourke amp Franks 1995)
5 Polydomy leads to increased nest densities Inpolygynous colonies polydomy reduces local mate
336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

336 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
competition (LMC) (ie brother-brother competitionfor mating) by increasing the number of females whichwill succeed in nest establishment locally (Pamilo ampRosengren 1984) thus reducing the bias towardsfemales However LMC has been only rarely shown tooccur in ants
Despite these predictions empirical evidence formale-biased sex ratios in polydomous colonies is weakand inconsistent the few existing studies givingstrongly contrasted results In two species sex ratioswere found to be even at the colony level(M punctiventris Snyder amp Herbers 1991 Tetrapon-era sp PSW 80 Buschinger et al 1994) Male-biasedsex ratios were found in polydomous colonies ofMyrmica rubra (Walin Seppauml amp Sundstroumlm 2001)as well as in several Formica species (RosengrenCherix amp Pamilo 1986 Sundstroumlm 1995) althoughfemale-biased sex ratios were found in populations ofFormica aquilonia (Pamilo amp Rosengren 1983) andF polyctena (R Rosengren amp M Elias unpubl data)However all five hypotheses described above considerthe colony level and are based directly or indirectlyupon the supposition that polydomous ant species arepolygynous Predictions under monogyny are morevague If workers win the conflict in queenless nestsfemale-biased sex ratios may be expected undermonogyny However if worker reproduction occursmale-biased sex ratios may arise in queenless nestsThus the question remains what are the effects of thenumber of queens on sex allocation in polydomous col-onies This question will be addressed in the followingtwo sections which compare patterns expected underpolygyny and monogyny
Among-nest variation in polygynous speciesIf several nests of a colony contain at least one queenthen within-colony-between-nest variation in sexratios and in particular sex ratio specialization at thenest level must be taken into account in explanationsof among-colony variation in sex ratios We presenthere these mechanisms as partially reviewed bySundstroumlm (1994) and discuss how each may beaffected by polydomy
1 Variations in resource levels may shift the nest sexratio towards a male or female bias
2 Colony- or nest-level sex allocation may varybecause of varying relative effects of competitionbetween males for females (LMC Hamilton 1967) andcompetition between females for resources nests andor food (local resource competition Clark 1978 Craig1980) due to differential dispersal of the sexes(inferred from various sex ratio patterns) HoweverLMC has remained controversial in ants mainlybecause (1) manipulation of sex allocation by workers
in response to relatedness asymmetries (Trivers ampHare 1976) is an additional powerful mechanism offemale bias and (2) the predominant mating systemin social insects is thought to make LMC unlikely(sexuals of most species mate randomly in nuptialflights Houmllldobler amp Wilson 1990) Nevertheless sev-eral species have evolved alternative mating tactics(limited male dispersal intranest copulation) andLMC has been shown to occur in some ants (Cremeramp Heinze 2002)
3 The smaller a nest the more it allocates to the sexthat does not experience resource competition becauseproductivity is more or less correlated with nest sizeThus large nests specialize in the other sex This maycause a highly biased colonypopulation sex ratio
4 Nests with a high relatedness asymmetry betweenworkers are predicted to specialize in the productionof the sex (generally females) to which they are rela-tively more related than is the average nest in thecolony Those with a lower relatedness asymmetry areselected to compensate any bias induced by the formerones and generally may specialize in males (Boomsmaamp Grafen 1991)
Thus peripheral nests of a colony are expected to beselected to produce long-distance dispersive femaleswhereas central nests should produce just a smallnumber of short-distance dispersive females andnumerous males dispersing both inside and outsidethe colony (Trivers amp Hare 1976 Nonacs 1986Bourke amp Franks 1995) A likely consequence is thatcentral nests recruit more females As proposed insome previous studies polygyny indirectly selects forlower queen longevity (Keller amp Genoud 1997 Keller1998) Queen turnover might thus be higher in centralthan in peripheral nests Relatedness asymmetrywould be greater in central than in peripheral nestssuch that central nests would produce more malesthan peripheral nests The runaway selection thuspredicted could be reduced if peripheral nests havegreater access to resources or if queens are activelyexchanged between nests leading to homogenizationof relatedness or of colony odour The balance betweendispersal strategy and relatedness asymmetry wouldthen affect the degree to which sex ratio is biased dif-ferentially between nests
If high population density leading to a significantincrease in the total amount of resources usable inthe inhabited patch leads also to food sharingbetween nests then the colony could bias its sex ratioIndeed in a polydomous colony some nests can bedependent on ants of other nests of the same colonyfor foraging This has been termed local resourceenhancement (LRE Schwarz 1988) This increase ofthe total amount of food available can be achieved by
POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 337
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
either active protection of the food sources (eg aggre-gations of aphids) from arthropod competitors preda-tors and parasitoids (through protection behavioursandor active exclusion from the local habitat of mostcompetitors of ants) or by more efficient use of theresources For example many aphid species used byants for honeydew can only survive if they are regu-larly lsquomilkedrsquo by attending ants and ants can enhancethe productivity of this food resource simply by con-suming it The colony thereby creates an lsquoartificialresource patchrsquo (Rosengren amp Pamilo 1983) Howeverresource enhancement by this mechanism is outpacedby nest multiplication within a limited patch Thereshould hence be a threshold point where saturation ofthe habitat (with ants and nests) exceeds LRE Thisthreshold point should depend on the size and pro-ductivity of the patch and it may never be reached inpractice if the patch size is lsquoinfinitely largersquo This maybe the case for example with some Formica speciesinhabiting vast forest areas A female bias can be pre-dicted below this threshold point and a male biasabove it for two reasons First queens are moreexpensive to produce than males The TriversndashWillard hypothesis would thus predict that a surplusof females is produced as long as the patch saturationremains under the threshold point (Trivers amp Wil-lard 1973) Second males are not only cheaper toproduce but are also in polygynous-polydomous colo-nies a better vehicle for genes to escape from the mis-ery of a degenerating resource patch because femalesin these cases often stay in the natal nest or dispersethrough budding However other authors haveargued that LRE would enhance the production ofsexuals overall rather than bias sex allocation(Herbers 1990)
In brief in polydomous and polygynous colonies sexallocation in different nests appears most likely to bedetermined by a combination of relatedness patternsand LRC However a colony- and a population-levelbias towards males is expected because of unavoid-able competition for resources between proximatenests and between queens for new nest sites near thenatal nest LRC may be the main factor that deter-mines sex ratio in polydomous colonies of polygynousant species
Among-nest variation in monogynous speciesThe main potential consequence of polydomy for allo-cation in monogynous ant species is a unique oppor-tunity for workers in queenless nests to evade queencontrol and thereby to sexualize brood Thus signifi-cant differences in sex allocation could be predictedbetween queenless and queenright nests Further-more the fact that nests tend to rear single-sex broodsshould generally reflect the intensity of the queenndashworker conflict although this could be partially an
arithmetical consequence of very small nest size inmany of the cases studied
Few studies have considered variation of allocationin polydomous but monogynous ant species and onlyone has clearly demonstrated that workers are striv-ing to evade queen control This study consideredM punctiventris (Myrmicinae) in which queenlessnests allocate more resources to reproductive femalesthan do queenright nests although all workersdescend from only one singly mated queen Here lsquosplitsex ratiosrsquo reflect chance expectations exacerbated bya biological cause polydomy which could be consid-ered to be the outcome of worker behaviours aimed attaking control over sex allocation (ie as a conse-quence of a strong queenndashworker conflict over invest-ment in producing males Snyder amp Herbers 1991Banschbach amp Herbers 1996b)
The four other studies of which we are aware con-sidered species in which workers seem not to escapequeen control The first treated Tetraponera sp PSW-80 near attenuata The colonies of this bamboo-nestingsouth-east Asian pseudomyrmecine ant are monogy-nous and highly polydomous (up to nine bamboo stemsinhabited) Gyne development is suppressed in thequeenrsquos internode and to a lesser extent in the inter-nodes of the same stem Despite the fact that gynesdevelop in queenless nests workers do not appear toescape queen control because the numerical and theinvestment sex ratios are 1 1 and 3 1 respectively(Buschinger et al 1994) as predicted by Trivers ampHare (1976) The same pattern was found in multinestcolonies of the monogynous ant species Cataglyphisiberica relatively more worker brood are present inthe queenright nest than in queenless nests and sex-ual brood is only found in queenless nests (CerdaDahbi amp Retana 2002) The third study concernsProtomognathus americanus (Myrmicinae) whosepolydomous colonies each contain a single queen thatis singly inseminated Worker reproduction has beenobserved in queenless and queenright nests andaccounts for more than 70 of all males Despite thisfact there is no evidence for a queenndashworker conflictMale-biased allocation ratios in this species are dueinstead to the effects of independent polydomous nestunits and LRC among queens (Foitzik amp Herbers2001) Finally in the facultatively polydomous butmonogynous species C mckeyi split sex ratios werefound whatever the colony structure (Debout et al2003) and workers do not appear to derive any benefitfrom polydomous structure in term of escape fromqueen control
VARIABILITY IN COLONY CLOSURE
Colony closure preserves the colonyrsquos social integrityand prevents intrusion by heterocolonial workers Col-
338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

338 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
ony closure depends on the capacities to recognize anddiscriminate between familiar individuals and unfa-miliar intruders which are usually attacked (Dahbiet al 1996) The claims made in several previousstudies that in polydomous ant species low intraco-lonial relatedness should result in inefficient nest-mate recognition are based on the assumption ofpolygyny not on polydomy per se (Keller amp Passera1989 Provost amp Cerdan 1990 Banschbach amp Her-bers 1996a)
The only studies on colony closure in polydomousants have been conducted on monogynous speciesCataglyphis iberica (Formicinae) and Cataulacusmckeyi (Myrmicinae) In the terricolous species Cata-glyphis iberica polydomy is not seasonal but allexchange of workers among nests is stopped in winterHowever colony closure remains very high and nest-mate recognition is unchanged after regrouping(Dahbi et al 1996) For this species colony integritymay be maintained through transport of young adultworkers (Dahbi et al 1997) and exchange of cuticularcompounds (Dahbi amp Lenoir 1998a) All workersshare the same cuticular hydrocarbon profile irre-spective of their provenance (queenright or queenlessnest) Colony odour in this polydomous and monogy-nous ant thus derives from worker cues and not onlyfrom the queen (Dahbi amp Lenoir 1998b) Similar con-clusions were reached concerning C mckeyi a tropicalplant-ant (Debout et al 2003) Both studies demon-strate that polydomy does not necessarily lead to arelaxation of colony closure even if rates of exchangeof workers between nests are very low
SPATIAL DISTRIBUTION OF NESTS WITHIN A POPULATION AND ITS CONSEQUENCES FOR GENETIC
STRUCTURE
Spatial aggregation of nests is the expected patternunder polydomy (Herbers 1985 1989) and it shouldlead to specific patterns of spatial genetic structureOne empirical study showed effectively that Lep-tothorax ambiguus (Myrmicinae) nests with similargenotypes tended to cluster in space generallyreflecting polydomous colonies (Herbers amp Grieco1994) Differentiation among subpopulations ofpolygynous polydomous populations of F truncorumhas also been detected contrasting with the lack ofdifferentiation among subpopulations of monogy-nous monodomous populations of the same species(Sundstroumlm 1993b) Recent studies on the polygy-nous polydomous ant F polyctena (Beye Neumannamp Moritz 1997) also support the idea of strong localgenetic structure as a result of the budding processassociated with polydomy (Elias et al 2002) Inpolydomous populations of Formica pratensis spa-tial and genetic distances were correlated again
indicating budding as the predominant dispersalmechanism (Beye et al 1998 Pirk et al 2001) Howthe genetic structure is related to the pattern of nestdistribution in such populations of polydomous colo-nies and how it affects competition and selection onsex ratios at the local scale are interesting and openquestions
A METHODOLOGY TO DESCRIBE POLYDOMY AND TO EXPLORE ITS EFFECTS
The boundaries of separate polydomous colonies in apopulation are often unknown There is a need forappropriate methods to permit the mapping of nests inpopulations of polydomous colonies following a stan-dardized methodology Colony boundaries can bedetermined according to spatial behavioural andgenetic criteria and preferably all of these in combi-nation Here we propose a step-by-step methodologyaimed at facilitating the distinction of new cases ofpolydomy so that the effects of polydomy on popula-tion structure and ecology can be more thoroughlyexplored The traits examined with the hypothesesjustifying their examination are compiled in Table 4
DESCRIPTION OF PARTICULAR SPATIAL PATTERNS
Nests can be mapped and analysed using the very effi-cient nearest-neighbour method (Clark amp Evans1954 Simberloff 1979) as in some cornerstone stud-ies such as those on Leptothorax spp (Herbers 1986)or on L neoniger (Traniello amp Levings 1986) Nestclustering is detected by calculating the distance tothe nearest neighbouring nest and comparing that totheoretical values calculated by simulation over allthe mapped nests
Table 4 Theoretical consequences of a polydomousstructure
(A) An aggregated spatial distribution of nests(B) Between-nest exchanges of individuals mainly work-
ers but also brood females and queens(C) Uniformity of workersrsquo chemical profiles all workers
of all the nests of a colony share one lsquogestaltrsquo odour(D) Strong colony closure vs low nest closure direct con-
sequence of points (B) and (C)(E) Colony male-biased sex-ratio a parsimonious expla-
nation is LRC (over nest sites) plus the constant-female hypothesis
(F) A strong genetic relationship between individuals of a same colony (regardless of nest provenance)
(G) Special patterns of abandonmentcolonization of nests depending on resource availability (seasonal polydomy)
POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 339
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
FOLLOWING MOVEMENTS OF INDIVIDUALS
Information on movements of workers or of dispersalof sexuals could be provided by direct observationsandor mass or individual marking of workers andorpupae followed by recapture (Rosengren amp Pamilo1983) However care must be taken in interpretingthe apparently cooperative nature of many of thenestndashnest interactions Reciprocal movements ofyoung workers and brood between nests often inter-preted as lsquocooperative exchanges between nestsrsquo mayalso be signs of competition or parasitism betweennests or attempts to alleviate overcrowding in theown nest while simultaneously flooding adjacent nestswith progeny genetically related to the carrier antsand their mothers Similarly workers visiting neigh-bouring nests may be stealing or begging food or couldeven be lsquospiesrsquo scouting a neighbouring nest before atake-over Likewise observations of trophallaxisbetween workers from different nests must be consid-ered with caution Studies using isotope-marked foodshowed actually that food exchange by trophallaxisbetween Formica aquilonia workers from distantnests occurs with the same high frequency as betweennestmate workers (Rosengren 1979) or more surpris-ingly that such food exchange even takes placebetween ants of different species (Bhatkar amp Kloft1977) Observation of lsquocooperationrsquo is thus not alonesufficient to establish the borders of polydomous colo-nies probably because trophallaxis is also an appease-ment signal correlated with at least mild aggression(R Rosengren pers comm)
DEGREE OF COLONY CLOSURE
Aggression tests conducted using pairwise reciprocalcomparisons of mature nests andor investigation ofvariability in the composition of the major cuticularhydrocarbons allow determination of the degree ofcolonynest closure (ie Dahbi et al 1996 Deboutet al 2003) Results of aggression experiments mustalso be interpreted with caution since the level ofaggression between workers from different nests (ofthe same or different colonies) may vary in space andtime and may depend on factors such as diet compo-sition or food availability (eg in L humile see Liangamp Silverman 2000 Silverman amp Liang 2001 Suarezet al 2002) In F polyctena R Rosengren also ob-served very rare cannibalistic lsquowarsrsquo within nests ofthe same colony Occurring in spring when activity re-sumes such battles among previously interconnectedand genetically related nests do not lead to rupture oftheir relations later on in the season Worker ex-change and reciprocal carrying of workers and broodcontinue as before the incident These spring battlesmay be due to severe starvation conditions (R Rosen-gren pers comm)
GENETIC ANALYSIS
Because of the limits of behavioural methods notedabove they must be complemented by genetic analy-ses to confirm the relatedness between individuals of aputative polydomous colony Whether occupants of dif-ferent nests belong to a single polydomous colony canbe assessed by comparing the relatedness of nestmateworkers (Rsame) with (1) their relatedness to workers ofthe nearest neighbour nest (Rnearest neighbour) deducedfrom aggression experiments to be concolonial or het-erocolonial and (2) their relatedness to workers of allother studied nests The overall frequency of polydomyamong nearest-neighbour nests in a population can beestimated by dividing Rnearest neighbour by Rsame (Pedersenamp Boomsma 1999) Methods of spatial correlationhierarchical F-statistics matrix correlation or regres-sion of genetic and spatial distance could also giveclues to detect similar worker genotypes among neigh-bour nests It should be noted that if nests containingonly one queen contain more than one matriline ofworkers three explanations are possible (1) thesemonogynous nests are fractions of larger polygynousand polydomous colonies (2) these nests have recentlylost a second queen whose worker offspring remain inthe nest and (3) there is considerable mixing of work-ers among colonies (lack of colony closure)
DETERMINATION OF SOCIAL STRUCTURE
Finally census of each caste for entire colonies (at thenest and entire colony levels) yields insights abouthow conflicts over resource and sex allocation areresolved within the colony and among its constituentnests The effects of polydomy will be most easilyexamined when there is variation in this trait withina single species because this situation provides theopportunity to relate this life-history trait to ecologicalfactors
WHAT REMAINS TO BE DISCOVERED ABOUT POLYDOMY IN ANTS
Much remains to be done on polydomy in ants We lackboth complete empirical studies and theoretical mod-els Here we present some suggestions for researchpriorities aimed at understanding in depth the phe-nomenon of polydomy and all its implications and atplacing this biological trait within the evolutionaryhistory of ants The main message in this review is topoint out the great need for adequate description ofthe life history traits of the studied species Generallythe biologies of particular ant species are known byonly a few individuals and it is very difficult to gainsatisfactory data on even simple traits such as thenumber of queens or the mean colony size One prior-ity is comparative study of different populations of
340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

340 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
species polymorphic for this trait or of closely relatedpolydomous and monodomous species Such studieswould enable distinction between those life-historytraits related to polydomy that are (proximate or ulti-mate) causal factors of polydomy and those that areadaptive traits related to polydomy or by-products ofit
Plant-ants or lsquophytoeciousrsquo ants (Quek et al 2004)namely ants that nests only in myrmecophytes (plantswith various usually specialized hollow stem- or leaf-cavities called domatia Davidson amp McKey 1993)may be particularly convenient models for studyingpolydomy for two reasons First boundaries of nests(and colonies) are clearly identifiable making experi-mentation easier than in many terricolous ants Sec-ond their polydomy may exist at several spatial scales(eg between and within trees) Indeed for all plant-ant species that we know in mature colonies all antson a single tree originate from a single colony Caneach domatium be considered a nest of a highly sub-divided colony Comparative studies of various antndashplant systems whose domatia differ in their architec-ture could allow the study of polydomous structures ata smaller spatial scale Among myrmecophytes theindividual plantrsquos population of domatia can be (1)spatially highly subdivided with each internode (egLeonardoxa McKey 1984) stipule-pair (eg AcaciaJanzen 1966) or leaf-pouch pair (eg Maieta Vascon-celos 1991) being a separate domatium (2) a systemintermediate in its degree of subdivision (eg Barteriafistulosa in which each long plagiotropic branch is aseparate domatium Janzen 1972) or (3) a singleinterconnected branching cavity extending through-out the plantrsquos stem system (eg Vitex lianas Djieacuteto-Lordon et al 2005)
Some scientists have noted the parallels betweenthe foraging strategies and modular growth of polyd-omous ant colonies and clonal plants (Harper 1977Harper amp Bell 1979) Workers of polydomous colonieslsquoforagersquo for new nest sites and subnests enable expan-sion of the area over which a colony collects foodresources such as ramets of a clonally growing plant(Loacutepez Serrano amp Acosta 1994) The application ofmodular demography theory (with its morphologicaland functional analysis of resource capture) to antecology must consider that an ant colony unlike clonalplants is never a single clone Conflicts between antsin a colony due to ever-present (even low) relatednessasymmetry will prevent total cooperation betweensubunits of a colony (Stevens 2000)
The existence of polydomy could aid in our under-standing of some within-colony social conflicts Forexample seasonal polydomy in a monogynous societymay provide opportunities to understand how queenndashworker conflicts are resolved However as we havealready noted empirical data are lacking even for
lsquowell-knownrsquo species For example here we onlyretained traits for which data were available for morethan 50 of species Other interesting traits related topolydomy could not be examined due to the fragmen-tary nature of data Even the mean number of nestsper colony cannot be compared among polydomousspecies No study has yet examined the relationshipsbetween worker polymorphism and polydomy Are dif-ferent worker morphs differentially distributed amongnests Worker monomorphism is assumed to be linkedto polygyny and colonies of taxa with polymorphicworkers are assumed to be less prone to be dispatchedover several nest sites (Frumhoff amp Ward 1992) How-ever in C gigas polydomy is associated with the exist-ence of a specialized lsquotransporterrsquo subcaste (Pfeiffer ampLinsenmair 1998) Data are also lacking on the differ-ence in queen longevity between mono- and poly-domous colonies within a species the existence ofeffective reproduction through budding and the relat-edness between nests
Finally we need to place polydomy into the contextof a global ant molecular phylogeny (for the most re-cently published ant molecular phylogeny see Moreauet al 2006) This is necessary to explore the origin ofpolydomy to determine how many times it appearedindependently and to examine associations betweenthe evolution of polydomy and that of polygyny
ACKNOWLEDGEMENTS
We thank A Dalecky M Dufayuml P Pamilo and P SWard for interesting discussions and valuablecomments on various drafts of the manuscript andespecially the late R Rosengren who thoroughly crit-icised commented on and annotated an almost finalversion of this review We are greatly indebted to thefollowing individuals for having kindly put their ownliterature databases and personal knowledge at ourdisposition C Djieto-Lordon M Kenne A Maeder JOrivel R Rosengren and P Stevens Our work onants has been supported by a thesis grant from theFrench Ministry of Education and Research (GD)and by grants from the Institut Franccedilais de la Biodi-versiteacute and from the National Geographic SocietyrsquosCommittee for Research and Exploration (DM andGD) ME acknowledges the financial support pro-vided through the European Community ImprovingHuman Potential Programme under contract HPRN-CT-2000minus00052 (INSECTS network) and under con-tract HPMF-CT-2002minus01781 (Marie Curie individualfellowship) The literature survey pertaining to thisstudy was concluded in January 2004 and was submit-ted in March 2005 This paper is dedicated to the lateR Rosengren who shared his irreplaceable knowledgeof Formica societies and who greatly improved thismanuscript
POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 341
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
REFERENCES
Ackonor JB 1981 The distribution of the ant Cataulacusguineensis F Smith (Hymenoptera Formicidae) in a Ghana-ian cocoa farm Insect Science and its Application 1 249ndash261
Ackonor JB 1983 The nest and nesting habits of the ant Cat-aulacus guineensis F Smith (Hymenoptera Formicidae) in aGhanaian coca farm Insect Science and its Application 4267ndash283
Acosta FJ Lopez F Serrano JM 1995 Dispersed versuscentral place foraging intra and intercolonial competition inthe strategy of trunk trail arrangement of a harvester antAmerican Naturalist 145 389ndash411
Adams ES 1990 Boundary disputes in the territorial antAzteca trigona effects of asymmetries in colony size AnimalBehaviour 39 321ndash328
Adams ES 1994 Territory defense by the ant Azteca trigonamaintenance of an arboreal ant mosaic Oecologia 97 202ndash208
Adams ES Levings SC 1987 Territory size and populationlimits in mangrove termites Journal of Animal Ecology 561069ndash1081
Alloway TM Buschinger A Talbot M Stuart RJ ThomasC 1982 Polygyny and polydomy in three North Americanspecies of the ant genus Leptothorax Mayr (HymenopteraFormicidae) Psyche 89 249ndash274
Anderson C McShea DW 2001 Intermediate-level parts ininsect societies adaptive structures that ants build awayfrom the nest Insectes Sociaux 48 291ndash301
Backus VL 1993 Packaging of offspring by nests of the antLeptothorax longispinosus parentndashoffspring conflict andqueen-worker conflict Oecologia 95 293ndash289
Backus VL 1995 Rules for allocation in a temperate forestant demography natural selection and queen-worker con-flict American Naturalist 145 775ndash796
Banschbach VS Herbers JM 1996a Complex colony struc-ture in social insects I Ecological determinants and geneticconsequences Evolution 50 285ndash297
Banschbach VS Herbers JM 1996b Complex colony struc-ture in social insects II Reproduction queen-worker con-flict and levels of selection Evolution 50 298ndash307
Banschbach VS Levit N Herbers JM 1997 Nest temper-atures and thermal preferences of a forest ant species is sea-sonal polydomy a thermoregulatory mechanism InsectesSociaux 44 109ndash122
Benzing DH 1991 Myrmecotrophy origins operation andimportance In Huxley CR Cutler DF eds Antndashplant inter-actions Oxford Oxford University Press 353ndash373
Beye M Neumann P Chapuisat M Pamilo P MoritzRFA 1998 Nestmate recognition and the genetic related-ness of nests in the ant Formica pratensis Behavioral Ecol-ogy and Sociobiology 43 67ndash72
Beye M Neumann P Moritz RFA 1997 Nestmate recogni-tion and the genetic gestalt in the mound-building ant For-mica polyctena Insectes Sociaux 44 49ndash58
Bhatkar AP Kloft WJ 1977 Evidence using radioactivephosphorus of interspecific food exchange in ants Nature265 140ndash142
Bhatkar AP Vinson SB 1987 Colony limits in Solenopsisinvicta Buren In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 599ndash600
Billick I 1999 The use of mark-recapture to measure workernumber in the rock nesting ant species Formica neorufibar-bis Emery Insectes Sociaux 46 256ndash260
Boomsma JJ Grafen A 1991 Colony-level sex ratio selec-tion in the eusocial Hymenoptera Journal of EvolutionaryBiology 3 383ndash407
Boomsma JJ van Loon AJ Brouwer AH Andrasfalvy A1990 Lasius neglectus a new polygynous pest ant in EuropeIn Veeresh GK Mallik B Viraktamath CA eds Socialinsects and the environment New Delhi IBH Publishers CoPvt Ltd 271ndash272
Bourke AFG Chan GL 1999 Queen-worker conflict oversexual production and colony maintenance in perennialsocial insects American Naturalist 154 417ndash426
Bourke AFG Franks NR 1995 Social evolution in antsPrinceton NJ Princeton University Press
Braun U Peeters C Houmllldobler B 1994 The giant nests ofthe African stink ant Paltothyreus tarsatus (Formicidae Pon-erinae) Biotropica 26 308ndash311
Bristow CM Cappaert D Campbell NJ Heise A 1992Nest structure and colony cycle of the Allegheny mound antFormica exsectoides Forel (Hymenoptera Formicidae)Insectes Sociaux 39 385ndash402
Bulmer MG 1983 Sex ratio theory in social insects withswarming Journal of Theoretical Biology 100 329ndash339
Bulmer MG Adams ES Traniello JFA 2001 Variation incolony structure in the subterranean termite Reticuliter-mes flavipes Behavioral Ecology and Sociobiology 49 236ndash243
Buschinger A Klein RW Maschwitz U 1994 Colony struc-ture of a bamboo-dwelling Tetraponera sp (HymenopteraFormicidae Pseudomyrmecinae) from Malaysia InsectesSociaux 41 29ndash41
Bustos X Cherix D 1998 Contribution agrave la biologie de Tap-inoma melanocephalum (Fabricius) (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 11 95ndash101
Carlin NF Reeve HK Cover SP 1993 Kin discriminationand division of labour among matrilines in the polygynouscarpenter ant Camponotus planatus In Keller L ed Queennumber sociality in insects Oxford Oxford University Press362ndash401
Cerda X Dahbi A Retana J 2002 Spatial patterns tempo-ral variability and the role of multi-nest colonies in a monog-ynous Spanish desert ant Ecological Entomology 27 7ndash15
Cerda X Retana J 1992 A behavioural study of transporterworkers in Cataglyphis iberica ant colonies (HymenopteraFormicidae) Ethology Ecology and Evolution 4 359ndash374
Cerda X Retana J de Haro A 1994 Social carryingbetween nests in polycalic colonies of the monogynous antCataglyphis iberica (Hymenoptera Formicidae) Sociobiol-ogy 23 215ndash231
Cerdan P Provost E 1990 Influence du nombre de reinesdans la fermeture de la socieacuteteacute chez Messor barbarus(Hym Formicidae) Actes Des Colloques Insectes Sociaux 6227ndash234
342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

342 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ceusters R 1979 Donneacutees deacutemographiques de diverses colo-nies polycaliques de Formica polyctena Foerst Lausanne CRIUSSI-FS 31ndash60
Chagne P Beugnon G Dejean A 2000 Fourragement chezGigantiops destructor (Fabricius) (Formicidae Formicinae)Actes Des Colloques Insectes Sociaux 13 21ndash26
Chapman RE Bourke FG 2001 The influence of socialityon the conservation biology of social insects Ecology Letters4 650ndash662
Cherix D 1986 Les fourmis des bois Lausanne Atlas VisuelsPayot
Cherix D 1987 Relation between diet and polyethism inFormica colonies Experientia Supplementum (Behavior inSocial Insects) 54 93ndash115
Cherix D Bourne JD 1980 A field study on a super-colonyof the red wood ants Formica lugubris Zett in relation toother predatory arthropods (spiders harvestmen and ants)Revue Suisse de Zoologie 87 955ndash973
Cherix D Maddalena-Feller C 1987 Foraging strategy inFormica bruni in relation to colony structure an importantstep towards polycalism In Eder J Rembold H eds Chem-istry and biology of social insects Munich Verlag J Peperny515ndash516
Cherix D Werner P Catzeflis F 1980 Organisation spa-tiale drsquoun systegraveme polycalique chez Formica (Coptoformica)exsecta Nyl (Hymenoptera Formicidae) Mitteilungen derSchweizerischen Entomologischen Gesellschaft 53 163ndash172
Clark AB 1978 Sex ratio and local resource competition in aprosimian primate Science 201 163ndash165
Clark PJ Evans FC 1954 Distance to nearest neighbour asa measure of spatial relationships in populations Ecology35 445ndash453
Clark DB Guayasamin C Pazmino O Donoso C Devil-lacis YP 1982 The tramp ant Wasmannia auropunctata minusautecology and effects on ant diversity and distribution onSanta-Cruz island Galapagos Biotropica 14 196ndash207
Collingwood CA 1987 Taxonomy and zoogeography of theFormica rufa L species group In Eder J Rembold H edsChemistry and biology of social insects Munich Verlag JPeperny 65ndash67
Conway JR 1996 A field study of the nesting ecology of thethatching ant Formica obscuripes Forel at high altitude inColorado Great Basin Naturalist 56 326ndash332
Conway JR 1997 Foraging activity trails food sources andpredators of Formica obscuripes Forel (Hymenoptera Formi-cidae) at high altitude in Colorado Pan-Pacific Entomologist73 172ndash183
Corn ML 1980 Polymorphism and polyethism in the neotro-pical ant Cephalotes atratus (L) Insectes Sociaux 27 29ndash42
Cosens D Toussaint N 1985 An experiment study of the for-aging strategy of the wood ant Formica aquilonia AnimalBehaviour 33 541ndash552
Craig R 1980 Sex investment ratios in social HymenopteraAmerican Naturalist 116 311ndash323
Cremer S Heinze J 2002 Adaptive production of fightermales queens of the ant Cardiocondyla adjust the sex ratiounder local mate competition Proceedings of the Royal Soci-ety of London Series B 269 417ndash422
Crozier RH Pamilo P 1996 Evolution of social insect colo-nies Oxford Oxford University Press
Crozier RH Pamilo P Crozier YC 1984 Relatedness andmicrogeographic genetic variation in Rhytidoponera mayrian Australian arid-zone ant Behavioral Ecology and Socio-biology 15 143ndash150
Curtis BA 1985 Nests of the Namib desert dune ant Cam-ponotus detritus Emery Insectes Sociaux 32 313ndash320
Cushman JH Rashbrook VK Beattie AJ 1994 Assessingbenefits to both participants in a lycaenidndashant associationEcology 75 1031ndash1041
Czechowski W 1990 A raid of Formica sanguinea Latr(Hymenoptera Formicidae) Przeglad Zoologiczny 19 449ndash463
Czechowski W 1999 Lasius fuliginosus (Latr) on a sandydune minus its living conditions and interference during raids ofFormica sanguinea Latr (Hymenoptera Formicidae)Annales Zoologici 49 117ndash123
Czechowski W Rotkiewicz W 1997 Relation between For-mica sanguinea Latr amp Formica cinerea cinerea Mayr(Hymenoptera Formicidae) minus an unusual form of dulosisAnnales Zoologici 47 469ndash478
Czechowski W Yamauchi K 1994 Intraspecific relations inCardiocondyla nuda (Mayr) (Hymenoptera Formicoidea)Memorabilia Zoologica 48 39ndash54
Dahbi A 1997 Reconnaissance et dynamique de lrsquoodeur colo-niale chez la fourmi Cataglyphis iberica PhD Thesis Uni-versiteacute Paris XIII
Dahbi A Cerda X Hefetz A Lenoir A 1996 Social closureaggressive behavior and cuticular hydrocarbon profiles inthe polydomous ant Cataplyphis iberica (Hymenoptera For-micidae) Journal of Chemical Ecology 122 2173ndash2186
Dahbi A Cerda X Hefetz A Lenoir A 1997 Adult trans-port in the ant Cataglyphis iberica a means to maintain auniform colonial odour in a species with multiple nestsPhysiological Entomology 22 13ndash19
Dahbi A Lenoir A 1998a Nest separation and the dynamicsof the Gestalt odor in the polydomous ant Cataglyphis iberica(Hymenoptera Formicidae) Behavioral Ecology and Socio-biology 42 349ndash355
Dahbi A Lenoir A 1998b Queen and colony odour in themultiple nest ant species Cataglyphis iberica (FormicidaeFormicinae) Insectes Sociaux 45 301ndash313
David CT Wood DL 1980 Orientation to tails by a carpen-ter ant Camponotus modoc (Hymenoptera Formicidae) ina giant sequoia forest Canadian Entomologist 112 993ndash1000
Davidson DW 1988 Ecological studies of neotropical ant gar-dens Ecology 69 1138ndash1152
Davidson DW 1997 The role of resource imbalances in theevolutionary ecology of tropical arboreal ants BiologicalJournal of the Linnean Society 61 153ndash181
Davidson DW McKey D 1993 The evolutionary ecology ofsymbiotic ant-plant relationships Journal of HymenopteraResearch 2 13ndash83
Dean WRJ 1989 Foraging and forager-recruitment in Oph-thalmopone hottentota Emery (Hymenoptera Formicidae)Psyche 96 123ndash130
POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 343
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Dean WRJ Yeaton RI 1993 The influence of harvester antMessor capensis nest-mounds on the productivity and distri-bution of some plant species in the southern Karoo southAfrica Vegetatio 106 21ndash35
Debout G Provost E Renucci M Tirard A Schatz BMcKey D 2003 Defining colony boundaries in a tropicalplant-ant behavioural chemical and genetical study of poly-domy in Cataulacus mckeyi (Myrmicinae) Oecologia 137195ndash204
DeHeer CJ Backus VL Herbers JM 2001 Sociogeneticresponses to ecological variation in the ant Myrmica punc-tiventris are context dependent Behavioral Ecology andSociobiology 49 375ndash386
Dejean A Akoa A Djieto-Lordon C Lenoir A 1994Mosaic ant territories in an African secondary rain forest(Hymenoptera Formicidae) Sociobiology 23 275ndash292
Dejean A Beugnon G Lachaud JP 1993 Spatial compo-nents of foraging behavior in an African ponerine ant Pal-tothyreus tarsatus Journal of Insect Behavior 6 271ndash285
Dejean A Djieto-Lordon C Durand JL 1997 Ant mosaicin oil palm plantations of the southwest province of Came-roon impact on leaf miner beetle (Coleoptera Chrysomel-idae) Journal of Economic Entomology 90 1092ndash1096
Dejean A Feacuteneacuteron R 1999 Predatory behaviour in the pon-erine ant Centromyrmex bequaerti a case of termitolestyBehavioral Processes 47 125ndash133
Dejean A Lachaud JP 1994 Ecology and behavior of theseed-eating ponerine ant Brachyponera senaarensis (Mayr)Insectes Sociaux 41 191ndash210
Dejean A McKey D Gibernau M Belin M 2000 The arbo-real ant mosaic in a Cameroonian rainforest (HymenopteraFormicidae) Sociobiology 35 403ndash423
Del Rio Pesado MG Alloway TM 1983 Polydomy in theslave-making ant Harpagoxenus americanus (Emery)(Hymenoptera Formicidae) Psyche 90 151ndash162
Delabie JHC 1995 Inquilinismo simultacircneo de duas espeacuteciesde Centromyrmex (Hymenoptera Formicidae Ponerinae) emcupinzeiros de Syntermes sp (Isoptera Termitidae Nasuter-mitinae) Revista Brasileira de Entomologia 39 605ndash609
Delabie JHC Benton FP de Medeiros MA 1991 La poly-domie chez les Formicidae arboricoles dans les cacaoyegraveres duBreacutesil optimisation de lrsquooccupation de lrsquoespace ou strateacutegiedeacutefensive Insectes Sociaux 7 173ndash178
Delage-Darchen B 1974 Ecologie et biologie de Cremato-gaster impressa Emery fourmi savanicole drsquoAfrique InsectesSociaux 21 13ndash34
Detrain C 1990 Field-study on foraging by the polymorphicant species Pheidole pallidula Insectes Sociaux 37 315ndash332
Djieacuteto-Lordon C Dejean A Ring RA Lauga J McKey D2005 Ecology of an improbable association the pseudomyr-mecine plant-ant Tetraponera tessmanni and the myrmeco-phytic vine Vitex thyrsiflora (Lamiaceae) in CameroonBiotropica 37 421ndash430
Elias M Rosengren R Sundstroumlm L 2005 Seasonal poly-domy and unicoloniality in a polygynous population of thered wood ant Formica truncorum Behavioral Ecology andSociobiology 57 339ndash349
Elias M Rosengren R Sundstroumlm L Pamilo P 2002Dynamics of the structure of a polydomous population of theant Formica polyctena Proceedings of the XIV InternationalCongress of IUSSI 27 July ndash 3 August 2002 Sapporo Hok-kaido University
Elmes GW 1987 Temporal variation in colony population ofthe ant Myrmica sulcinodis I Changes in queen numberworker number and spring production Journal of AnimalEcology 56 559ndash571
Espadeler X Rey S 2001 Biological constraints and colonyfounding in the polygynous invasive ant Lasius neglectus(Hymenoptera Formicidae) Insectes Sociaux 48 159ndash164
Federle W Maschwitz U Fiala B 1998 The two-partnerant-plant system of Camponotus (Colobopsis) sp1 andMacaranga puncticulata (Euphorbiaceae) natural history ofthe exceptional ant partner Insectes Sociaux 45 1ndash16
Fernandez-Escudero I Seppauml P Pamilo P 2001 Depen-dent colony founding in the ant Proformica longisetaInsectes Sociaux 48 80ndash82
Fisher R 1938 Statistical methods for research workers 10thedn Edinburgh Oliver and Boyd Press
Fletcher DJC Blum MS Whitt TV Temple N 1980Monogyny and polygyny in the fire ant Solenopsis invictaAnnals of the Entomological Society of America 73 658ndash661
Foitzik S Herbers JM 2001 Colony structure of a slave-making ant I Intracolony relatedness worker reproductionand polydomy Evolution 55 307ndash315
Forel A 1874 Les fourmis de la Suisse Systeacutematique noticesanatomiques et physiologiques architecture distributiongeacuteographique nouvelles expeacuteriences et observations demoeurs Neue Denkschriften der Allgemeinen Schweiz-erischen Gesellschaft der Gesammten Naturwissenschaften26 1ndash452
Francoeur A Peacutepin D 1978 Productiviteacute de la fourmi For-mica dakotensis dans la pessiegravere tourbeuse 2 Variationsannuelles de la densiteacute des colonies de lrsquooccupation des nidset de la reacutepartition spatiale Insectes Sociaux 25 13ndash30
Frank SA 1998 Foundations of social evolution PrincetonNJ Princeton University Press
Frumhoff PC Ward PS 1992 Individual-level selection col-ony-level selection and the association between polygynyand worker monomorphism in ants American Naturalist139 559ndash590
Gadau J Gertsch P Heinze J Pamilo P Houmllldobler B1998 Oligogyny by unrelated queens in the carpenter antCamponotus ligniperdus Behavioral Ecology and Sociobiol-ogy 44 23ndash33
Gerst H 2001 Testing the hypothesis of polydomy in the sub-alpine ant Formica podzolica Boulder CO University ofColorado at Boulder Honors Program Available at httpucsubcoloradoedu~honorsthesess01gersthtml
Gibernau M Dejean A 2001 Ant protection of a het-eropteran trophobiont against a parasitoid wasp Oecologia126 53ndash57
Giraud T Pedersen JS Keller L 2002 Evolution of super-colonies the Argentine ants of southern Europe Proceedingsof the National Academy of Science of the United States ofAmerica 99 6075ndash6079
344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

344 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Greenslade PJM Halliday RB 1983 Colony dispersion andrelationships of meat ants Iridomyrmex purpureus and alliesin an arid locality in south Australia Insectes Sociaux 3082ndash99
Hamilton WD 1967 Extraordinary sex ratios Science 156477ndash488
Hannonen M Sundstroumlm L 2003 Sociobiology worker nep-otism among polygynous ants Nature 421 910
Harkness RD Isham V 1988 Relations between nests ofMessor wasmanni in Greece Insectes Sociaux 35 1ndash18
Harper JL 1977 Population biology of plants London Aca-demic Press
Harper JL Bell AD 1979 The population dynamics ofgrowth form in organisms with modular construction InAnderson RM Turner BD Taylor LR eds Populationdynamics 20th Symposium of the British Ecological SocietyOxford Blackwell Scientific Publications 29ndash53
Hasegawa E 1992 Annual life cycle and timing of male-eggproduction in the ant Colobopsis nipponicus (Wheeler)Insectes Sociaux 39 439ndash446
Heinze J Foitzik S Hippert A Houmllldobler B 1996 Appar-ent dear-enemy phenomenon and environment-based recog-nition cues in the ant Leptothorax nylanderi Ethology 102510ndash522
Helms KR 1999 Colony sex ratios conflict between queensand workers and apparent queen control in the ant Pheidoledesertorum Evolution 53 1470ndash1478
Helms KR Fewell J Rissing S 2000 Sex ratio determina-tion by queens and workers in the ant Animal Behaviour 59523ndash527
Herbers JM 1984 Social organization in Leptothorax antswithin- and between-species patterns Psyche 90 361ndash386
Herbers JM 1985 Seasonal structuring of a north temperateant community Insectes Sociaux 32 224ndash240
Herbers JM 1986 Nest site limitation and facultative polyg-yny in the ant Leptothorax longispinosus Behavioral Ecol-ogy and Sociobiology 19 115ndash122
Herbers JM 1987 Population structure in Leptothorax long-ispinosus In Eder J Rembold H eds Chemistry and biologyof social insects Munich Verlag J Peperny 35ndash36
Herbers JM 1989 Community structure in north temperateants temporal and spatial variation Oecologia 81 201ndash211
Herbers JM 1990 Reproduction investment and allocationratios for the ant Leptothorax longispinosus sorting out thevariation American Naturalist 136 178ndash208
Herbers JM 1991 The population biology of Tapinomaminutum (Hymenoptera Formicidae) in Australia InsectesSociaux 38 195ndash204
Herbers JM DeHeer CJ Foitzik S 2001 Conflict over sexallocation drives conflict over reproductive allocation inperennial social insect colonies American Naturalist 158178ndash192
Herbers JM Grieco S 1994 Population structure of Lep-tothorax ambiguus a facultatively polygynous and polydo-mous ant species Journal of Evolutionary Biology 7 581ndash598
Herbers JM Tucker CW 1986 Population fluidity in Lep-tothorax longispinosus (Hymenoptera Formicidae) Psyche93 217ndash229
Higashi S 1979 Polygyny nest budding and internest mix-ture of individuals in Formica (Serviformica) japonica Mots-chulsky at Ishikari shore Kontyucirc 47 381ndash389
Hoffmann BD 1998 The big-headed ant Pheidole megaceph-ala a new threat to monsoonal northwestern AustraliaPacific Conservation Biology 4 250ndash255
Houmllldobler B 1983 Territorial behavior in the green tree ant(Oecophylla smaragdina) Biotropica 15 241ndash250
Houmllldobler B 1984 Communication during foraging andnest-relocation in the African stink ant Paltothyreus tarsa-tus Fabr (Hymenoptera Formicidae Ponerinae) Zeitschriftfuumlr Tierpsychologie 65 40ndash52
Houmllldobler B Lumsden CJ 1980 Territorial strategies inants Science 210 732ndash739
Houmllldobler B Moumlglich M 1980 The foraging system ofPheidole militicida (Hymenoptera Formicidae) InsectesSociaux 27 237ndash264
Houmllldobler B Wilson EO 1977 The number of queens animportant trait in ant evolution Naturwissenschaften 64 8ndash15
Houmllldobler B Wilson EO 1986 Ecology and behaviorof the primitive cryptobiotic ant Prionopelta amabilis(Hymenoptera Formicidae) Insectes Sociaux 33 45ndash58
Houmllldobler B Wilson EO 1990 The ants Cambridge MABelknap Press
Holway DA Case TJ 2000 Mechanisms of dispersed cen-tral-place foraging in polydomous colonies of the Argentineant Animal Behaviour 59 433ndash441
Ichinose K 1987 Annual life cycle of Paratrechina flavipes(Hymenoptera Formicidae) in the Tomakomai experimentforest southern Hokkaido Kontyucirc 54 9ndash20
Ito F Higashi S Maeta Y 1988 Growth and development ofCamponotus (Paramyrmamblys) kiusiuensis Santschi colo-nies (Hym Formicidae) Insectes Sociaux 35 251ndash261
Janzen DH 1966 Coevolution of mutualism between antsand acacias in Central America Evolution 20 249ndash275
Janzen DH 1972 Protection of Barteria (Passifloraceae) byPachysima ants (Pseudomyrmecinae) in a Nigerian rain for-est Ecology 53 885ndash892
Janzen DH 1973 Evolution of polygynous obligate acacia-ants in western Mexico Journal of Animal Ecology 42 727ndash750
Jayasuriya AK Traniello JFA 1985 The biology of theprimitive ant Aneuretus simoni (Emery) (FormicidaeAneuretinae) I Distribution abundance colony structureand foraging ecology Insectes Sociaux 32 363ndash374
Johnson RN Crozier RH 1998 Population viscosity andmulti-nest colonies in the ant Polyrhachis doddi In Sved Jed Genetics Society of Australia 45th annual conference Syd-ney Genetics Society of Australia 33
Kannowski PB 1959 The use of radioactive phosphorus inthe study of colony distribution of the ant Lasius minutusEcology 40 162ndash165
Kaspari M Vargo EL 1995 Colony size as a buffer againstseasonality Bergmannrsquos rule in social insects AmericanNaturalist 145 610ndash632
Keller L ed 1993 Queen number sociality in insects OxfordOxford University Press
POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 345
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Keller L 1991 Queen number mode of colony founding andqueen reproductive success in ants (Hymenoptera Formi-cidae) Ethology Ecology and Evolution 3 307ndash316
Keller L 1995 Social life the paradox of multiple queen col-onies Trends in Ecology and Evolution 10 355ndash360
Keller L 1998 Queen lifespan and colony characteristics inants and termites Insectes Sociaux 45 235ndash246
Keller L Genoud M 1997 Extraordinary lifespans in ants atest of evolutionary theories of ageing Nature 389 958ndash960
Keller L Passera L 1989 Influence of the number of queenson nestmate recognition and attractiveness of queens toworker in the Argentine ant Iridomyrmex humilis (Mayr)Animal Behaviour 37 733ndash740
Keller L Passera L 1990 Fecundity of ant queens in relationto their age and the mode of colony founding InsectesSociaux 37 116ndash130
Kenne M 1999 Biologie drsquoune fourmi terricole drsquoimportanceeacuteconomique Myrmicaria opaciventris Emery (HymenopteraMyrmicinae) PhD Thesis Universiteacute Paul Sabatier minusToulouse III
Kenne M Dejean A 1999 Spatial distribution size and den-sity of nests of Myrmicaria opaciventris Emery InsectesSociaux 46 179ndash185
Klein RW 1987 Colony structure of three species ofPseudomyrmex (Hymenoptera Formicidae Pseudomyrmeci-nae) in Florida In Eder J Rembold H eds Chemistry andbiology of social insects Munich Verlag J Peperny 107ndash108
Klotz JH Reid BL Klotz SA 1996 Trailing the elusive car-penter ant a key to its control American Entomology Spring1996 33ndash39
Le Masne G 1994 Remarques sur lrsquoeacutevolution du comporte-ment des myrmeacutecophiles Memorabilia Zoologica 48 115ndash132
Leston D 1978 A neotropical ant mosaic Annals of the Ento-mological Society of America 71 649ndash653
Leacutevieux J 1983 Mode drsquoexploitation des ressources alimen-taires eacutepigeacutees des savanes africaines par la fourmi Myrmi-caria eumenoides Gerstaecker Insectes Sociaux 30 165ndash176
Leacutevieux J Diomande T 1978 La nutrition des fourmisgranivores I Cycle drsquoactiviteacute et reacutegime alimentaire deMessor galla et de Messor (= Craptomyrmex) regalis(Hymenoptera Formicidae) Insectes Sociaux 25 127ndash139
Levings SC Traniello JFA 1981 Territoriality nest dis-persion and community structure in ants Psyche 88 265ndash319
Liang D Silverman J 2000 lsquoYou are what you eatrsquo diet mod-ifies cuticular hydrocarbons and nestmate recognition in theArgentine ant Linepithema humile Naturwissenschaften 87412ndash416
Liefke C Dorow WHO Houmllldobler B Maschwitz U 1998Nesting and food resources of syntopic species of the antgenus Polyrhachis (Hymenoptera formicidae) in West-Malaysia Insectes Sociaux 45 411ndash425
Longino JT 1991 Azteca ants in Cecropia trees taxonomycolony structure and behaviour In Huxley C Cutler D edsAntndashplant interactions Oxford Oxford University Press271ndash287
Longino JT 2000 Ants in Costa Rica [on line] evergreenOlympia WA State College Available at httpwwwevergreeneduantsAntsofCostaRicahtml
Loacutepez F Serrano JM Acosta FJ 1994 Parallels betweenthe foraging strategies of ants and plants Trends in Ecologyand Evolution 9 150ndash153
Mabelis A 1994 Flying as a survival strategy for wood ants ina fragmented landscape (Hymenoptera Formicidae) Memo-rabilia Zoologica 48 147ndash170
Macevicz S 1979 Some consequences of Fisherrsquos sex ratioprinciple for social Hymenoptera that reproduce by colonyfission American Naturalist 113 363ndash371
MacKay WP MacKay EE 1984 Why do harvester ants storeseeds within nests Sociobiology 9 31ndash47
Maeder A Cherix D 2001 Problegravemes poseacutes par Formicaparalugubris une nouvelle espegravece de fourmis des bois ActesDes Colloques Insectes Sociaux 14 21ndash25
Majer JD 1976 The ant mosaic in Ghana cocoa farms fur-ther structural considerations Journal of Applied Ecology13 145ndash155
Maschwitz U Moog J 2000 Communal peeing a newmode of flood control in ants Naturwissenschaften 87563ndash565
McGlynn TP 1999 Non-native ants are smaller than relatednative ants American Naturalist 154 690ndash699
McIver JD 1991 Dispersed central place foraging in Austra-lian meat ants Insectes Sociaux 38 129ndash137
McIver JD Steen T 1994 Use of a secondary nest in GreatBasin desert thatch ants (Formica obscuripes Forel) GreatBasin Naturalist 54 359ndash365
McKey D 1984 Interaction of the ant-plant Leonardoxa afri-cana (Caesalpiniaceae) with its obligate inhabitants in arainforest in Cameroon Biotropica 16 81ndash99
Mercier JL Dejean A 1996 Ritualized behavior during com-petition for food between two Formicinae Insectes Sociaux43 17ndash29
Mercier JL Lenoir A Dejean A 1994 Polydomoussocieties of the tree-dwelling ant Polyrhachis laboriosa(F Smith) In Lenoir A Arnold G Lepage M eds Lesinsectes sociaux 12th congress of the International Unionfor the Study of Social Insects Paris Publications Univer-isteacute Paris-Nord 472
Morais HC 1994 Coordinated group ambush a new preda-tory behavior in Azteca ants (Dolichoderinae) InsectesSociaux 41 339ndash342
Moreau CS Bell CD Vila R Archibald SB Pierce NB2006 Phylogeny of ants diversification in the age ofangiosperms Science 312 101ndash104
Nickerson JC Cromroy HL Whitcomb WH Cornell JA1975 Colony organization and queen numbers in two speciesof Conomyrma Annals of the Entomological Society of Amer-ica 68 1083ndash1085
Nonacs P 1986 Ant reproductive strategies and sex alloca-tion theory Quarterly Review of Biology 61 1ndash21
Nonacs P 1993 Polygyny colony life history and opti-mal sex investment In Keller L ed Queen number andsociality in insects Oxford Oxford University Press110ndash131
346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

346 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
OrsquoNeil KM 1988 Trail patterns and movement of workersamong nests in the ant Formica obscuripes (HymenopteraFormicidae) Psyche 95 1ndash13
Orivel J 2000 Lrsquoadaptation agrave la vie arboricole de la fourmiPachycondyla goeldii (Hymenoptera Ponerinae) PhD The-sis Universiteacute Paris XIII
Orivel J Errard C Dejean A 1997 Ant gardens interspe-cific recognition in parabiotic ant species Behavioral Ecologyand Sociobiology 40 87ndash93
Pamilo P 1991 Life span of queens in the ant Formicaexsecta Insectes Sociaux 38 111ndash119
Pamilo P Crozier RH Fraser J 1985 Inter-nest interac-tions nest autonomy and reproductive specialization in anAustralian arid-zone ant Rhytidoponera sp12 Psyche 92217ndash236
Pamilo P Rosengren R 1983 Sex ratio strategies in For-mica ants Oikos 40 24ndash35
Pamilo P Rosengren R 1984 Evolution of nesting strate-gies of ants genetic evidence from Formica ants BiologicalJournal of the Linnean Society 21 331ndash348
Pamilo P Sundstroumlm L Fortelius W Rosengren R 1994Diploid males and colony-level selection in Formica antsEthology Ecology and Evolution 6 221ndash235
Partridge LW Partridge KA Franks NR 1997 Field sur-vey of a monogynous leptothoracine ant evidence of seasonalpolydomy Insectes Sociaux 44 75ndash83
Passera L 1993 Quels sont les caractegraveres eacutetho-physiologiques des lsquofourmis vagabondesrsquo Actes Des Col-loques Insectes Sociaux 8 39ndash45
Passera L 1994 Characteristics of tramp ants In WilliamsD ed Exotic ants Boulder CO Westview 23ndash43
Passera L Gilbert M Aron S 2001 Social parasitism inants effects of the inquiline parasite Plagiolepis xene St onqueen distribution and worker production of its host Plagi-olepis pygmaea Latr Insectes Sociaux 48 74ndash49
Pedersen JS Boomsma JJ 1999 Genetic analyses of colonystructure in polydomous and polygynous ant populationsBiological Journal of the Linnean Society 66 115ndash144
Peeters C 1993 Monogyny and polygyny in ponerine antswith or without queens In Keller L ed Queen number andsociality in insects Oxford Oxford University Press 234ndash261
Peeters C Crewe R 1986 Male biology in the queenless pon-erine ant Ophthalmopone berthoudi (Hymenoptera Formi-cidae) Psyche 93 277ndash284
Peng RK Christian K Gibb K 1998 Locating queenant nests in the green ant Oecophylla smaragdina(Hymenoptera Formicidae) Insectes Sociaux 45 477ndash480
Perfecto I 1994 Foraging behavior as a determinant of asym-metric competitive interaction between two ant species in atropical agroecosystem Oecologia 98 184ndash192
Pfeiffer M Linsenmair KE 1998 Polydomy and the orga-nization of foraging in a colony of the Malaysian giant antCamponotus gigas (Hymenoptera Formicidae) Oecologia117 579ndash590
Pfeiffer M Linsenmair KE 2000 Contributions to the lifehistory of the Malaysian giant ant Camponotus gigas(Formicidae) Insectes Sociaux 47 123ndash132
Pfeiffer M Linsenmair KE 2001 Territoriality in theMalaysian giant ant Camponotus gigas (HymenopteraFormicidae) Journal of Ethology 19 75ndash85
Pirk CWW Neumann P Moritz RFA Pamilo P 2001Intranest relatedness and nestmate recognition in themeadow ant Formica pratensis (R) Behavioral Ecology andSociobiology 49 366ndash374
Pisarski B Czechowski W 1990 Modaliteacutes de colonisationdes fourmis du groupe Formica rufa au parc national deGorce (Pologne) Actes des Colloques Insectes Sociaux (ParisFrance) 6 237ndash242
Provost E Cerdan P 1990 Experimental polygyny and col-ony closure in the ant Messor barbarus (L) (Hym Formi-cidae) Animal Behaviour 115 114ndash126
Punttila P 1996 Succession forest fragmentation and thedistribution of wood ants Oikos 75 291ndash298
Quek SP Davies SJ Itino T Pierce NE 2004 Codiversi-fication in an ant-plant mutualism stem texture and theevolution of host use in Crematogaster (Formicidae Myrmic-inae) inhabitants of Macaranga (Euphorbiaceae) Evolution58 554ndash570
Reuter M Balloux F Lehmann L Keller L 2001 Kinstructure and queen execution in the Argentine ant Linepi-thema humile Journal of Evolutionary Biology 14 954ndash958
Reuter M Keller L 2001 Sex ratio conflict and worker pro-duction in eusocial Hymenoptera American Naturalist 158166ndash177
Roisin Y Pasteels JM Braekman JC 1986 Systegravemes poly-caliques chez Nasutitermes princeps (Desneux) Actes DesColloques Insectes Sociaux 3 123ndash132
Rosengren R 1979 The labial gland syndrome as an epi-demic in a polycalic group of nests of the wood ant speciesFormica aquilonia (Hymenoptera Formicidae) MemorandaSocietatis Pro Fauna Flora Fennica 55 73ndash84
Rosengren R 1986 Competition and coexistence in aninsular ant community a manipulation experiment(Hymenoptera Formicidae) Annales Zoologici Fennici 23297ndash302
Rosengren R Cherix D Pamilo P 1985 Insular ecologyof the red wood ant Formica truncorum Fabr I Polydo-mous nesting population size and foraging Mitteilungender Schweizerischen Entomologischen Gesellschaft 58 147ndash175
Rosengren R Cherix D Pamilo P 1986 Insular ecology ofthe red wood ant Formica truncorum Fabr II Distributionreproductive strategy and competition Mitteilungen derSchweizerischen Entomologischen Gesellschaft 59 63ndash94
Rosengren R Pamilo P 1983 The evolution of polygyny andpolydomy in mound-building Formica ants Acta Entomolog-ica Fennicca 42 65ndash77
Ross KG Fletcher DJC 1985 Comparative study of geneticand social structure in two forms of the fire ant Solenopsisinvicta (Hymenoptera Formicidae) Behavioral Ecology andSociobiology 17 349ndash356
Rowe HC Bristow M 1999 Sex ratio and sexual dimor-phism in Formica exsectoides the Allegheny mound ant(Hymenoptera Formicidae) Great Lakes Entomologist 32207ndash218
POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

POLYDOMY IN ANTS 347
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
Ruumlppell O Heinze J 1999 Alternative reproductive tacticsin females the case of size polymorphism in winged antqueens Insectes Sociaux 46 6ndash17
Sanders NJ Gordon DM 2000 The effects of interspecificinteractions on resource use and behavior in a desert antOecologia 125 436ndash443
SAS Institute Inc 1999 SAS statistical software release802 Cary NC SAS Institute Inc
Sasaki K Satoh T Obara Y 1996 Cooperative foundation ofcolonies by unrelated foundresses in the ant Polyrhachismoesta Insectes Sociaux 43 217ndash226
Savolainen R Vepsaumllaumlinen K 1988 A competition hierar-chy among boreal ants impact on resource partitioning andcommunity structure Oikos 51 135ndash155
Savolainen R Vepsaumllaumlinen K 1989 Niche differentiation ofant species within territories of the wood ant Formicapolyctena Oikos 56 3ndash16
Savolainen R Vepsaumllaumlinen K Deslippe RJ 1996 Repro-ductive strategy of the slave ant Formica podzolica relativeto raiding efficiency of enslaver species Insectes Sociaux 43201ndash210
Schmid-Hempel P 1987 Foraging characteristics of thedesert ant Cataglyphis Experientia Supplementum 54 43ndash61
Schwarz MP 1988 Local resource enhancement and sex-ratios in a primitively social bee Nature 331 346ndash348
Seifert B 2000 Rapid range expansion in Lasius neglec-tus (Hymenoptera Formicidae) minus an Asian invaderswamps Europe Mitteilungen Aus Dem Museum fuumlrNaturkunde Berlin Deutsche Entomologische Zeitschrift47 173ndash179
Silverman J Liang D 2001 Colony disassociation followingdiet partition in a unicolonial ant Naturwissenschaften 8873ndash77
Simberloff DS 1979 Nearest neighbour assessments of spa-tial configurations of circles rather than points Ecology 60679ndash685
Smith-Glaser TA 1994 RAPDs as a tool to discern polydomyin Formica pallidefulva nitidiventris In Lenoir A Arnold GLepage M eds Les Insectes sociaux 12th Congress of the Inter-national Union for the Study of Social Insects Paris Univer-siteacute Paris Nord 522
Snyder LE Herbers JM 1991 Polydomy and sexual alloca-tion ratios in the ant Myrmica punctiventris BehavioralEcology and Sociobiology 28 409ndash415
Stevens P 2000 The ecology of polygyny and polydomy inants introductory research essay no 23 Helsinki Univer-sity of Helsinki
Stuart RJ 1985 Spontaneous polydomy in laboratorycolonies of the ant Leptothorax curvispinosus Mayr(Hymenoptera Formicidae) Psyche 92 71ndash81
Stuart RJ 1987 Transient nestmate recognition cues contrib-ute to a multicolonial population structure in the ant Lep-tothorax curvispinosus Behavioral Ecology and Sociobiology21 229ndash235
Stuart RJ 1991 Nestmate recognition in leptothoracine antstesting for effects of queen number colony size and species ofintruder Animal Behaviour 42 277ndash284
Suarez AV Holway DA Liang D Tsutsui ND Case TJ2002 Spatiotemporal patterns of intraspecific aggression inthe invasive Argentine ant Animal Behaviour 64 697ndash708
Sundstroumlm L 1989 Genetic relatedness and populationstructure in Formica truncorum Fabr (Hymenoptera Formi-cidae) Actes Des Colloques Insectes Sociaux 5 93ndash100
Sundstroumlm L 1993a Foraging responses of Formica trun-corum (Hymenoptera Formicidae) minus exploiting stable vsspatially and temporally variable resources Insectes Sociaux40 147ndash161
Sundstroumlm L 1993b Genetic population structure and socio-genetic organisation in Formica truncorum (HymenopteraFormicidae) Behavioral Ecology and Sociobiology 33 345ndash354
Sundstroumlm L 1994 Sex ratio bias relatedness asymmetryand queen mating frequency in ants Nature 367 266ndash268
Sundstroumlm L 1995 Sex allocation and colony maintenance inmonogyne and polygyne colonies of Formica truncorum(Hymenoptera Formicidae) the impact of kinship and mat-ing structure American Naturalist 146 182ndash201
Torossian C 1960 La biologie de la fourmi Dolichoderusquadripunctatus (Hymeacutenoptegravere Formicoidea Dolichode-ridae) Insectes Sociaux 7 383ndash393
Torossian C 1974 Biologie et eacutethologie drsquoun ergatan-dromorphe de Dolichoderus quadripunctatus (L)(Hymenoptera Formicoidea Dolichoderidae) InsectesSociaux 21 145ndash150
Traniello JFA 1982 Population structure and social organi-zation in the primitive ant Amblyopone pallipes(Hymenoptera Formicidae) Psyche 89 65ndash80
Traniello JFA 1989 Chemical trail systems orientation andterritorial interactions in the ant Lasius neoniger Journal ofInsect Behaviour 2 339ndash353
Traniello JFA Levings SC 1986 Intra- and intercolony pat-terns of nest dispersion in the ant Lasius neoniger correla-tions with territoriality and foraging ecology Oecologia 69413ndash419
Trivers RL Hare H 1976 Haplodiploidy and the evolution ofthe social insects Science 191 249ndash263
Trivers RL Willard DE 1973 Natural selection of parentalability to vary sex-ratio of offspring Science 179 90ndash92
Tsuji K 1988 Nest relocation in the Japanese queenless antPristomyrmex pungens Mayr (Hymenoptera Formicidae)Insectes Sociaux 35 321ndash340
Tsuji K Furukawa T Kinomura K Takamine HYamauchi K 1991 The caste system of the dolichoderineant Technomyrmex albipes (Hymenoptera Formicidae) mor-phological description of queens workers and reproductivelyactive intercastes Insectes Sociaux 38 413ndash422
Tsuji K Yamauchi K 1994 Colony level sex allocation in apolygynous and polydomous ant Behavioral Ecology andSociobiology 34 157ndash167
Tsutsui ND Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepithema humile) in itsnative and introduced ranges Evolution 55 976ndash985
Tsutsui ND Suarez AV Holway DA Case TJ 2000Reduced genetic variation and the success of an invasive spe-
348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292

348 G DEBOUT ET AL
copy 2007 The Linnean Society of London Biological Journal of the Linnean Society 2007 90 319ndash348
cies Proceedings of the National Academy of Science of theUnited States of America 97 5948ndash5953
Vasconcelos HL 1991 Mutualism between Maieta guianen-sis Aubl a myrmecophytic melastome and one of its antinhabitants ant protection against insect herbivores Oeco-logia 87 295ndash298
Vasconcelos HL Davidson DW 2000 Relationship betweenplant size and ant associates in two Amazonian ant-plantsBiotropica 32 100ndash111
Vepsaumllaumlinen K Savolainen R Tiainen J Vilen J 2000Successional changes of ant assemblages from virgin andditched bogs to forests Annales Zoologici Fenniciici 37 135ndash149
Wagner D 1997 The influence of ant nests on Acacia seedproduction herbivory and soil nutrients Journal of Ecology85 83ndash93
Wagner D 2000 Pollen viability reduction as a potential costof ant association for Acacia constricta (Fabaceae) AmericanJournal of Botany 87 711ndash715
Walin L Seppauml P Sundstroumlm L 2001 Reproductive alloca-tion within a polygyne polydomous colony of the ant Myr-mica rubra Ecological Entomology 26 537ndash546
Walker J Stamps J 1986 A test of optimal caste ratio theoryusing the ant Camponotus (Colobopsis) impressus Ecology67 1052ndash1062
Waloff N Blackith RE 1962 The growth and distribution ofthe mounds of Lasius flavus (Fabricius) (Hym Formicidae) inSilwood Park Berkshire Journal of Animal Ecology 31 421ndash437
Way MJ 1954 Studies of the life history and ecology of the antOecophylla longinoda Latreille Bulletin of EntomologicalResearch 45 93ndash112
Weber NA 1935 The biology of the thatching ant Formicarufa obscuripes Forel in North Dakota Ecological Mono-graphs 5 165ndash206
Wellenstein G 1929 Die Ameisenkolonie Neue Beobachtun-gen an Ameisen Die Umschau 33 754ndash757
Wilson EO 1953 The origin and evolution of polymorphism inants Quarterly Review of Biology 28 136ndash156
Yamauchi K Kimura Y Corbara B Kinomura K Tsuji K1996 Dimorphic ergatoid males and their reproductivebehavior in the ponerine ant Hypoponera bondroiti InsectesSociaux 43 119ndash130
Yamauchi K Yoshida T Ogawa T Itoh S Ogawa YJimbo S Imai HT 2001 Spermatogenesis of diploid malesin the formicine ant Lasius sakagamii Insectes Sociaux 4828ndash32
Zakharov AA 1994 Primary and secondary federations inants Memorabilia Zoologica 48 279ndash292







