HOME RANGE, HABITAT USE, AND NEST SITE CHARACTERISTICS OF MISSISSIPPI KITES IN THE WHITE RIVER NATIONAL WILDLIFE REFUGE, ARKANSAS TROY J. BADER 1,2,3 AND JAMES C. BEDNARZ 1 ABSTRACT.—We located 39 Mississippi Kite (Ictinia mississippiensis) nests during the 2004 and 2005 breeding seasons in the White River National Wildlife Refuge in the Mississippi Alluvial Valley of Arkansas. Radio transmitters were placed on seven adult and eight juvenile kites; 649 locations based on ground telemetry and 393 locations from aircraft were recorded for six adults (n 5 5 males, 1 female) and five juveniles (n 5 2 males, 3 females). The mean 90% kernel home range was 3,098 ha for adult kites (n 5 6) and 439 ha (n 5 5) for juveniles. Radio-marked Mississippi Kites significantly used mature forest (65.6%), second growth (15.5%), and water (10.0%) relative to availability (59.6, 11.5, and 7.1%, respectively). Agriculture fields (6.5%) and wetlands (1.5%) were used significantly less within the home ranges of kites relative to availability (17.6 and 3.9%, respectively). Tree height, diameter at breast height (dbh), and height of nest tree emergence above the surrounding trees were significantly greater for nest sites than randomly-selected overstory trees. Nest trees were significantly closer to the edge than randomly-selected trees. Most (57%) Mississippi Kite nests were in crotches of secondary branches in the nest tree. Conservation of super-emergent trees and mature forests is needed for nesting and foraging areas. Second-growth forest should be allowed to mature (.75 years) and uniform canopies should be avoided. Received 7 August 2009. Accepted 28 April 2010. The Mississippi Kite (Ictinia mississippiensis) breeds in bottomland and riparian areas through- out the Mississippi River Valley and southeastern North America (e.g., Kalla 1979, Evans 1981, Whitmar 1987, Barber 1995, St. Pierre 2006). This raptor also inhabits riparian areas, shelter- belts, oak (Quercus spp.)-shrub prairies, and savannas in the western United States (Parker 1988). This kite has adapted to nesting in urban golf courses, parks, and neighborhoods primarily in the western portion of its range (Parker 1988). The ability of the western population to adapt to encroachment of humans has allowed it to increase in number and expand into new areas (Parker and Ogden 1979). Kite populations in the eastern United States were affected by severe habitat alteration in the late 1800s and early 1900s. Gosselink and Lee (1989) estimated that, at time of European settlement, there were ,80 million ha of forested freshwater wetlands in the United States with the majority in the Mississippi Alluvial Valley. The area of bottomland hardwood forests in the Mississippi Alluvial Valley decreased to 4.8 million ha by 1937, and to only 2.1 million ha by 1970 (Gosselink et al. 1990). Several researchers have described habitat use and nest site characteristics of Mississippi Kites, often based on small sample sizes, in Tennessee (Kalla 1979), Illinois (Evans 1981), Missouri (Whitmar 1987, Barber 1995), and Arkansas (St. Pierre 2006) within the Mississippi Alluvial Valley; almost all of this work remains unpublished. Allan and Sime (1943) observed daily activities of 169 Mississippi Kites in the panhandle of Texas and subjectively estimated home-range size to be ,250 ha. Barber (1995) using telemetry estimated mean home-range size of kites along the Mis- sissippi River in Missouri as 1,160 ha (n 5 10), much larger than estimates from the Texas panhandle. Barber (1995) compared the number of telemetry locations within specific vegetation types to estimates of the proportions of these types available in the surrounding landscape. Forested habitats fragmented with agricultural fields and developments are not ecologically equivalent to the relatively contiguous-forested habitats once used by kites in the 1800s and early 1900s. The vast changes in the landscape probably affected home-range size and habitat use, and may have influenced reproductive success of this species (Bader and Bednarz 2009). A better understanding of how Mississippi Kites use landscapes that are present today will be useful to conserve and enhance habitats for this species. Our objectives were to examine: (1) use of habitats and landscapes, and (2) selection of nest sites by Mississippi Kites in the Mississippi Alluvial Valley. The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:00 706 Cust # 09-121 1 Department of Biological Sciences, Arkansas State University, P. O. Box 599, Jonesboro, AR 72467, USA. 2 Current address: USDA-ARS, Stuttgart National Aqua- culture Research Center, P. O. Box 1050, Stuttgart, AR 72160, USA. 3 Corresponding author; e-mail: [email protected]The Wilson Journal of Ornithology 122(4):706–715, 2010 706
Transcript
HOME RANGE, HABITAT USE, AND NEST SITE CHARACTERISTICS
OF MISSISSIPPI KITES IN THE WHITE RIVERNATIONAL WILDLIFE REFUGE, ARKANSAS
TROY J. BADER1,2,3 AND JAMES C. BEDNARZ1
ABSTRACT.—We located 39 Mississippi Kite (Ictinia mississippiensis) nests during the 2004 and 2005 breeding
seasons in the White River National Wildlife Refuge in the Mississippi Alluvial Valley of Arkansas. Radio transmitters
were placed on seven adult and eight juvenile kites; 649 locations based on ground telemetry and 393 locations from
aircraft were recorded for six adults (n 5 5 males, 1 female) and five juveniles (n 5 2 males, 3 females). The mean 90%
kernel home range was 3,098 ha for adult kites (n 5 6) and 439 ha (n 5 5) for juveniles. Radio-marked Mississippi Kites
significantly used mature forest (65.6%), second growth (15.5%), and water (10.0%) relative to availability (59.6, 11.5, and
7.1%, respectively). Agriculture fields (6.5%) and wetlands (1.5%) were used significantly less within the home ranges of
kites relative to availability (17.6 and 3.9%, respectively). Tree height, diameter at breast height (dbh), and height of nest
tree emergence above the surrounding trees were significantly greater for nest sites than randomly-selected overstory trees.
Nest trees were significantly closer to the edge than randomly-selected trees. Most (57%) Mississippi Kite nests were in
crotches of secondary branches in the nest tree. Conservation of super-emergent trees and mature forests is needed for
nesting and foraging areas. Second-growth forest should be allowed to mature (.75 years) and uniform canopies should be
avoided. Received 7 August 2009. Accepted 28 April 2010.
The Mississippi Kite (Ictinia mississippiensis)
breeds in bottomland and riparian areas through-
out the Mississippi River Valley and southeastern
North America (e.g., Kalla 1979, Evans 1981,
Whitmar 1987, Barber 1995, St. Pierre 2006).
This raptor also inhabits riparian areas, shelter-
belts, oak (Quercus spp.)-shrub prairies, and
savannas in the western United States (Parker
1988). This kite has adapted to nesting in urban
golf courses, parks, and neighborhoods primarily
in the western portion of its range (Parker 1988).
The ability of the western population to adapt to
encroachment of humans has allowed it to
increase in number and expand into new areas
(Parker and Ogden 1979).
Kite populations in the eastern United States
were affected by severe habitat alteration in the
late 1800s and early 1900s. Gosselink and Lee
(1989) estimated that, at time of European
settlement, there were ,80 million ha of forested
freshwater wetlands in the United States with the
majority in the Mississippi Alluvial Valley. The
area of bottomland hardwood forests in the
Mississippi Alluvial Valley decreased to 4.8
million ha by 1937, and to only 2.1 million ha
by 1970 (Gosselink et al. 1990).
Several researchers have described habitat useand nest site characteristics of Mississippi Kites,often based on small sample sizes, in Tennessee(Kalla 1979), Illinois (Evans 1981), Missouri(Whitmar 1987, Barber 1995), and Arkansas (St.Pierre 2006) within the Mississippi Alluvial Valley;almost all of this work remains unpublished. Allanand Sime (1943) observed daily activities of 169Mississippi Kites in the panhandle of Texas andsubjectively estimated home-range size to be,250 ha. Barber (1995) using telemetry estimatedmean home-range size of kites along the Mis-sissippi River in Missouri as 1,160 ha (n 5 10),much larger than estimates from the Texaspanhandle. Barber (1995) compared the numberof telemetry locations within specific vegetationtypes to estimates of the proportions of these typesavailable in the surrounding landscape.
Forested habitats fragmented with agriculturalfields and developments are not ecologicallyequivalent to the relatively contiguous-forestedhabitats once used by kites in the 1800s and early1900s. The vast changes in the landscape probablyaffected home-range size and habitat use, and mayhave influenced reproductive success of thisspecies (Bader and Bednarz 2009). A betterunderstanding of how Mississippi Kites uselandscapes that are present today will be usefulto conserve and enhance habitats for this species.Our objectives were to examine: (1) use ofhabitats and landscapes, and (2) selection of nestsites by Mississippi Kites in the MississippiAlluvial Valley.
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:00 706 Cust # 09-121
1 Department of Biological Sciences, Arkansas State
University, P. O. Box 599, Jonesboro, AR 72467, USA.2 Current address: USDA-ARS, Stuttgart National Aqua-
culture Research Center, P. O. Box 1050, Stuttgart, AR
The Wilson Journal of Ornithology 122(4):706–715, 2010
706
METHODS
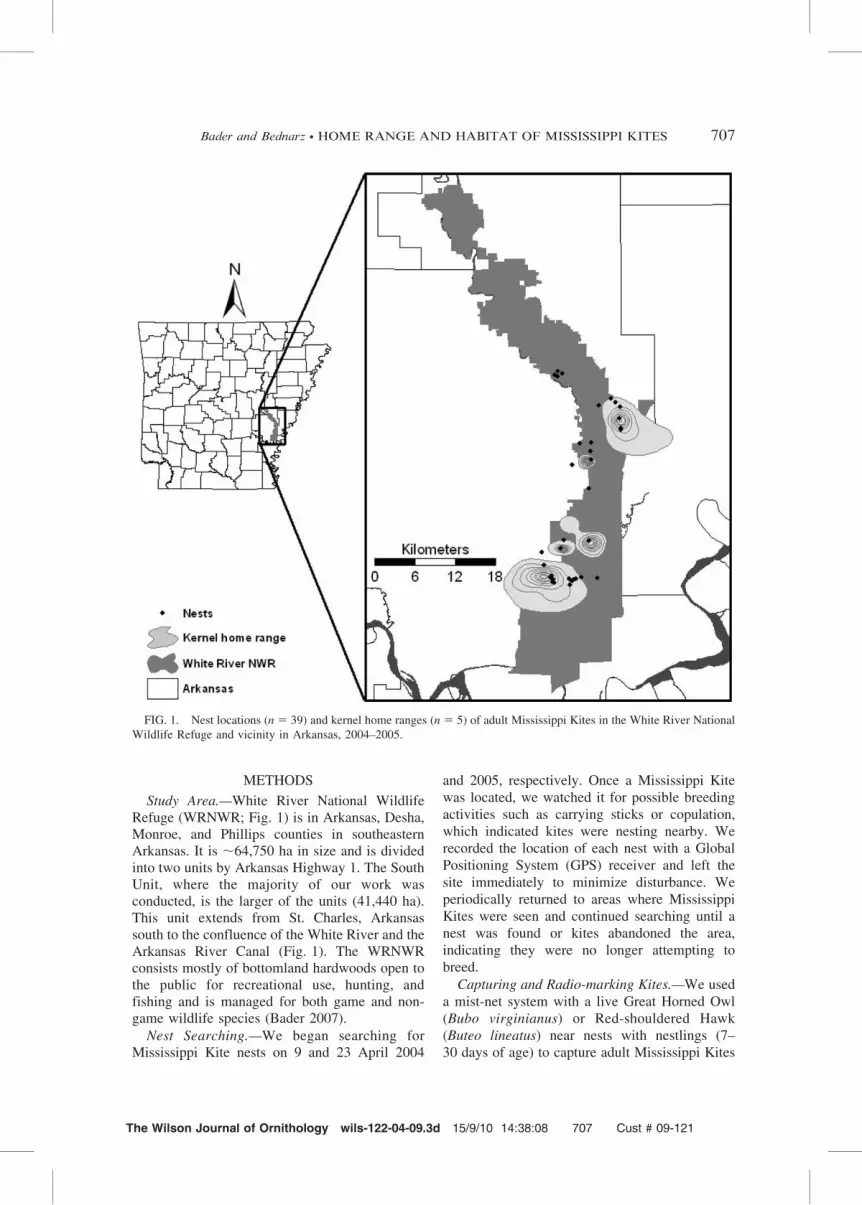
Study Area.—White River National WildlifeRefuge (WRNWR; Fig. 1) is in Arkansas, Desha,Monroe, and Phillips counties in southeasternArkansas. It is ,64,750 ha in size and is dividedinto two units by Arkansas Highway 1. The SouthUnit, where the majority of our work wasconducted, is the larger of the units (41,440 ha).This unit extends from St. Charles, Arkansassouth to the confluence of the White River and theArkansas River Canal (Fig. 1). The WRNWRconsists mostly of bottomland hardwoods open tothe public for recreational use, hunting, andfishing and is managed for both game and non-game wildlife species (Bader 2007).
Nest Searching.—We began searching forMississippi Kite nests on 9 and 23 April 2004
and 2005, respectively. Once a Mississippi Kite
was located, we watched it for possible breeding
activities such as carrying sticks or copulation,
which indicated kites were nesting nearby. We
recorded the location of each nest with a Global
Positioning System (GPS) receiver and left the
site immediately to minimize disturbance. We
periodically returned to areas where Mississippi
Kites were seen and continued searching until a
nest was found or kites abandoned the area,
indicating they were no longer attempting to
breed.
Capturing and Radio-marking Kites.—We used
a mist-net system with a live Great Horned Owl
(Bubo virginianus) or Red-shouldered Hawk
(Buteo lineatus) near nests with nestlings (7–
30 days of age) to capture adult Mississippi Kites
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:08 707 Cust # 09-121
FIG. 1. Nest locations (n 5 39) and kernel home ranges (n 5 5) of adult Mississippi Kites in the White River National
Wildlife Refuge and vicinity in Arkansas, 2004–2005.
Bader and Bednarz N HOME RANGE AND HABITAT OF MISSISSIPPI KITES 707
(Hamerstrom 1963, Barber et al. 1998). The mist-net system consisted of two 2.6 3 6 m mist nets(72 mm mesh) connected to a pulley system ontelescoping metal poles. This allowed the top ofthe net to be elevated ,7 m above ground withthe bottom of the lower net ,1.5 m off theground. The total dimensions of the system usingtwo mist nets were 5.2 m in height 3 6 m wide(Bader 2007).
We placed a U.S. Geological Survey (USGS)aluminum band and two or three plastic colorbands on legs of each kite for unique identifica-tion. Selected kites were also fitted with a radiotransmitter using a modified figure-eight legharness (Radley 2002). Mississippi Kites werefitted with a 6.0-g transmitter (Holohil SystemsLtd., Carp, ON, Canada). We recorded severallinear measurements, mass, and obtained a bloodsample from each kite, before it was released.Gender of each kite were assigned based onplumage characteristics (Parker 1999), and veri-fied by DNA analysis in the laboratory (Bader2007). We left the nest site immediately once akite was released to minimize any potential stresson adult and nestling kites.
We climbed to each nest and banded thenestling(s) when they were 3–4 weeks of age. Themass of each nestling was measured, and a USGSaluminum band and two or three plastic color bandswere placed on their legs for identification ofindividuals. Each nestling was also fitted with aradio transmitter using the procedure described foradults and returned to the nest.
Gender Identification.—A 0.05-cc sample ofblood was drawn from the brachial vein of eachkite using a hypodermic needle and a smallcapillary tube (Faaborg et al. 1995) and placed inLongmire Solution (Longmire et al. 1988). Theprotocol outlined by Donohue and Dufty (2006)was followed to extract DNA from the blood andisolate the CHD-Z and CHD-W bands (Griffiths etal. 1998) to identify the gender of captured kites.
Radiotelemetry of Adult and Juvenile Kites.—Triangulation was used to obtain locations fromthe ground and a fixed-wing aircraft was used toidentify point locations from the air. Receiversites for triangulations were established ,500 mapart along roads or all-terrain vehicle trails nearnest sites and compass azimuths were recorded inthe direction of the strongest signal from theradio-marked kite. Compass azimuths from atleast three receiver sites were taken within a 5-min period to estimate the error ellipse around
each location. Eight triangulations with a $10-min interval between each triangulation werecompleted 2–3 days each week. The 10-minperiod allowed time for the kite to move betweentriangulation attempts, minimizing autocorrelationbetween locations (Swihart and Slade 1985). Thetriangulation azimuth was entered into the OTATriangulation Program (Ripper et al. 2007), whichprovided coordinates for the estimated location.We could calculate an error ellipse using thisprogram that theoretically has a 95% probabilityof including the true location of the kite. Weeliminated all estimated locations that had anerror ellipse greater than 10 ha to improveaccuracy. This relatively large error ellipsecriteria was chosen to accommodate the highmobility of kites (typical home range size.3,000 ha).
We obtained locations from the air (DeVault etal. 2003) for each radio-marked kite 2–3 times perflight depending on the amount of flight timeavailable. Locations of individual kites wererecorded .20 min apart. Flights were conducted2–3 days each week using a Cessna 172 or 182equipped with a pair of ‘‘side-looking’’ 4-elementyagi antennae (Bader 2007).
Habitat Use.—Point locations were entered intoGeographic Information System (GIS) ArcViewsoftware and home-range polygons were devel-oped for each kite. Two types of home-rangepolygons were generated using point locations;minimum convex polygon (MCP) and 90% kernel(White and Garrott 1990). We used the 90%probability of use contours as estimates of home-range size. The home-range polygon of each kitewas overlaid onto USGS Digital OrthophotographQuarter Quadrangles (DOQQs) taken in fall 2004.
ArcMap 8.3 at a scale of 1:3,000 was used toclassify habitats into seven cover types withineach home range. The mature forest cover typeconsisted of tracts of large trees that had relativelyeven canopy with few openings and had not beenrecently logged (.30 year). Second-growth covertype consisted of young forest with large openingsin the canopy, abandoned fields, and recently-logged forest. The water cover type included allopen waterways (e.g., lakes, bayous, and rivers).The wetland cover type included all forested andnon-forested swamps and low-lying vernal areas.The agriculture cover type included all types ofagriculture fields (e.g., crop and hay fields). Theurban cover type consisted of noticeably devel-oped areas (e.g., rural towns). The road cover type
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:13 708 Cust # 09-121
708 THE WILSON JOURNAL OF ORNITHOLOGY N Vol. 122, No. 4, December 2010
included highways, county roads, and obviousprivate roads. Polygons were drawn around eachcover type and the sum of all polygons within acover type was used to calculate the number ofhectares available of that cover type within agiven home range.
Bailey’s confidence intervals (Bailey 1980,Cherry 1996) were used to examine whether kitesused cover types within their home range inproportion with that available. We used the Chi-square distribution and a Bonferroni correctionbased on the number of available cover types toconstruct a 95% confidence interval (CI) aroundthe proportion of telemetry locations within eachcover type. The confidence interval was comparedto the proportion of habitat that was availablewithin the kite’s home range to delineate the levelof kite use.
Nest-site Characteristics.—Vegetation datawere collected at every nest site after youngfledged or the nest failed and at an equal numberof sites selected using a random-number table.The distance (0–250 m) was paced while using acompass to follow the random azimuth to locatethe reference sample site and the closest overstorytree was chosen as the plot center. The samevegetation data were collected within a 0.04-hacircular plot around nest and random trees. Wefollowed the BBIRD Protocol (Martin et al. 1997)and included species of the tree, tree height, treediameter at breast height (dbh), distance to forestedge, dominant and co-dominant trees within thesurrounding area, and species and height of eachtree in the overstory and within the circular plot.Data collected on nest-site characteristics includ-ed nest height, orientation of nest in tree, andposition of nest in tree (primary or secondaryfork).
We collected data on perch trees, defined as thetree(s) in which kites perched in the morning andfrequently during the day in the immediatevicinity (,200 m) of their nests. Measurementscollected at perch trees were the same as thosetaken for nest trees, but also included distancefrom nest and azimuth to nest. We recordedwhether the tree was dead or alive and classified itas a primary or secondary perch tree. The primaryperch tree was the tree that one or both kites usedthe majority (.50%) of the time. The secondaryperch tree, if present, was the tree used by one orboth kites less frequently (,50%).
Statistical Analysis.—We used a paired t-test(SAS Institute 1999) to compare Mississippi Kite
nest sites with sites selected at random (Sokal andRohlf 1995), and a t-test to compare characteris-tics of primary and secondary perch trees.
RESULTS
We located 21 Mississippi Kite nests in 2004and 18 in 2005. We captured seven adult kites,each from separate nests (4 in 2004, 3 in 2005), in22 trapping attempts. We made 11 attempts tocapture chicks in 2004 and 2005 and capturednine (at ages of ,3.5–4 weeks) from nine nests.
Analysis of DNA to identify gender of capturedadult and nestling kites was conducted on 14blood samples (6 adults and 8 nestlings). Fivemale and one female adult kites, and four maleand four female nestling kites were identified.Five of six adults (83%) were identified correctlyto gender based on plumage coloration.
Radio-marked Mississippi Kites were locatedusing ground receiver sites from 9 July to 28August 2004 and from 14 July to 20 August 2005.Locations of each radio-marked kite were record-ed from a fixed-wing aircraft from 28 July to 19August 2004 and from 20 July to 26 August 2005.We recorded 649 locations from the ground (20045 449 locations, 2005 5 200 locations) and 393locations from the air (2004 5 134 locations,2005 5 259 locations).
MCP and 90% kernel home ranges weregenerated for each radio-marked kite (Table 1,Fig. 2). The mean (6 SE) MCP home range sizefor adult male radio-marked kites was 3,289.1 6
1,419.3 ha; n 5 5). Home-range data werecollected for only one female and the MCP homerange was 1,899.8 ha. The mean (6 SE) MCPhome range for male juvenile kites was 536.2 6
10.8 ha; n 5 2) and 508.4 6 316.4 ha; n 5 3) forfemale juvenile kites. The mean (6 SE) homerange using 90% kernel method for adult maleradio-marked kites was 3,567.3 6 1,393.5 ha; n 5
5). The home range size for the one radio-markedadult female kite using the 90% kernel methodwas 752.2 ha. The mean 90% kernel home rangefor male juvenile kites was 299.0 6 23.0 ha (n 5
2) and 533.0 6 266.7 ha (n 5 3) for femalejuvenile kites.
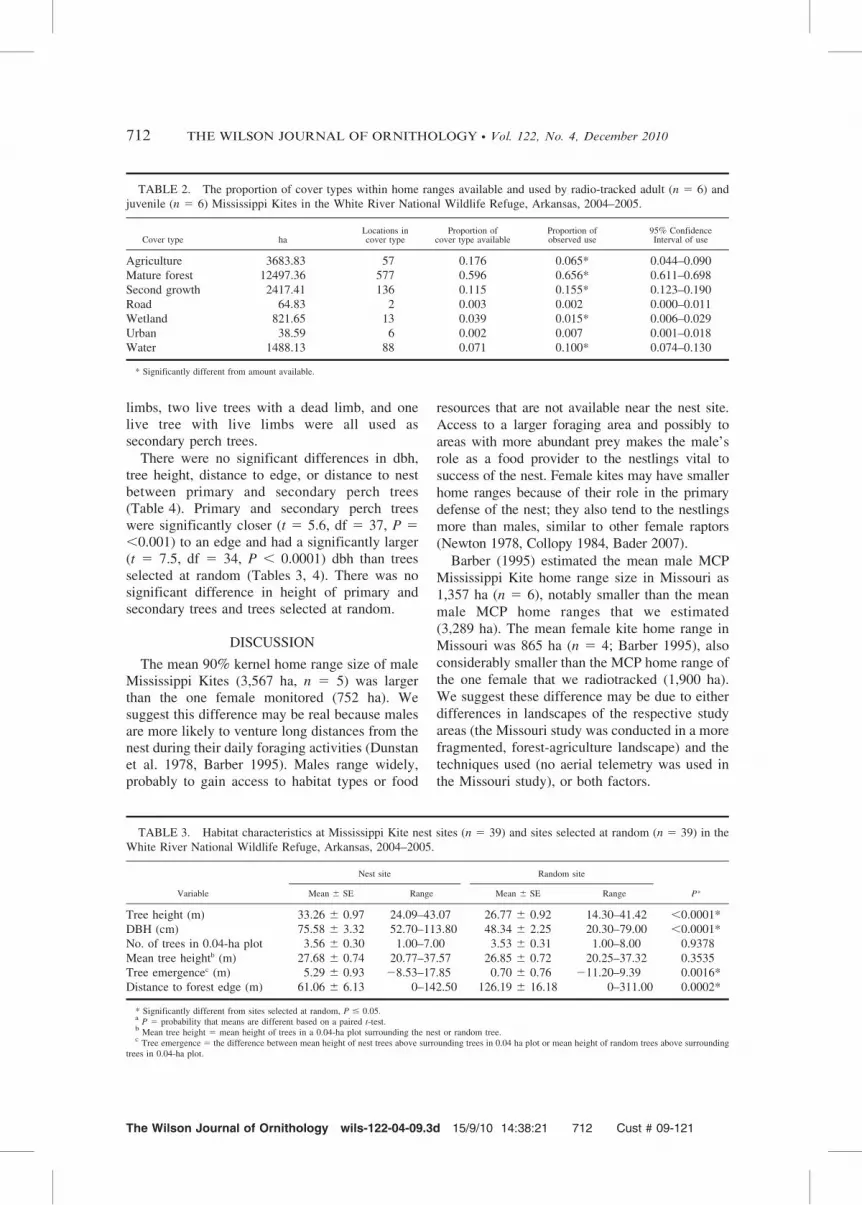
We estimated habitat use within home ranges of12 Mississippi Kites (6 adults, 6 juveniles).Mississippi Kites significantly used mature forest(95% CI 5 0.611–0.698; Table 2) more than itsavailability (0.596). Water was also significantlyused (95% CI 5 0.074–0.130; Table 2) morerelative to its availability (0.071). Mississippi
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:13 709 Cust # 09-121
Bader and Bednarz N HOME RANGE AND HABITAT OF MISSISSIPPI KITES 709
Kites also significantly used second growth (95%CI 5 0.123–0.190; Table 2) more relative to itsavailability (0.115). Agriculture fields and wet-lands were significantly used less relative toavailability (Table 2). Mississippi Kites used bothroads and urban areas in proportion to theiravailability (Table 2).
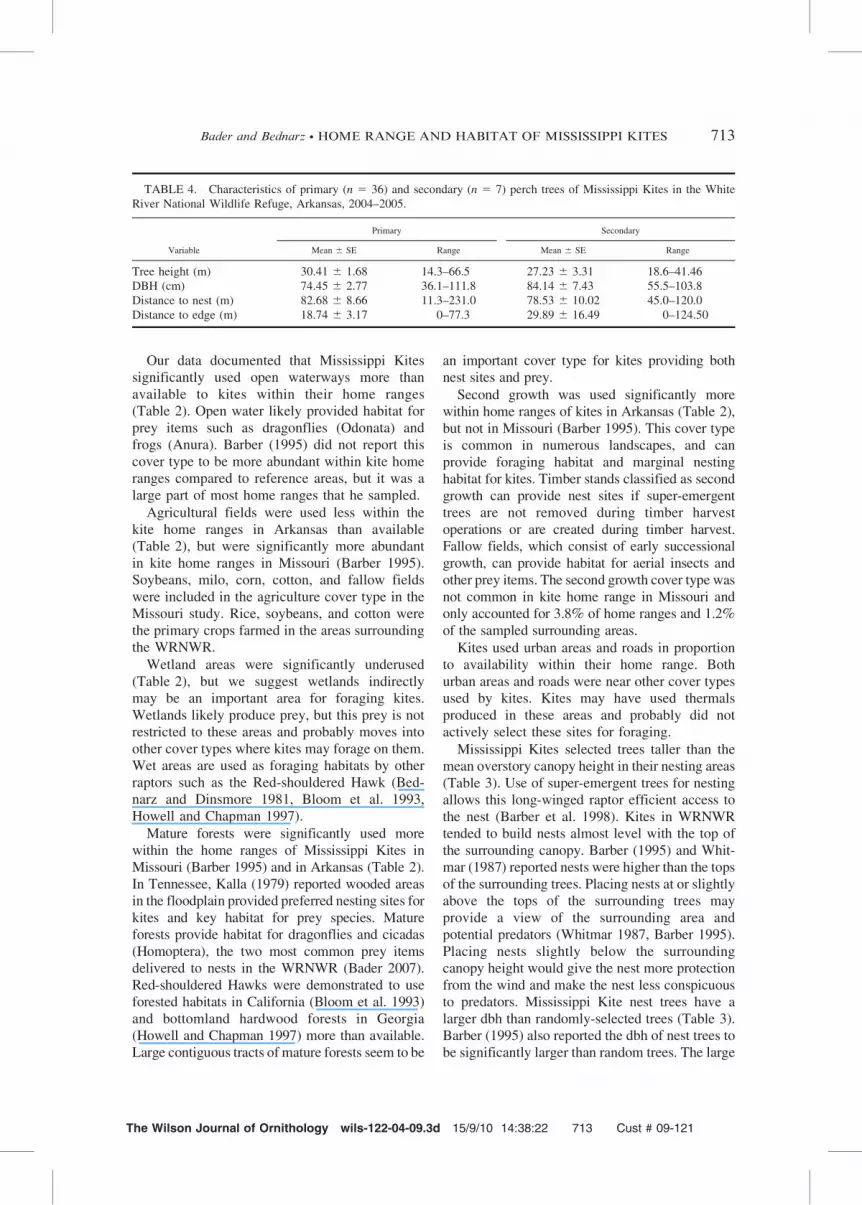
The mean nest tree height (33.26 m) wassignificantly taller (t 5 5.37, df 5 33, P ,
0.0001; Table 3) than paired randomly-selectedtrees (26.77 m). Mean dbh of nest trees (75.58 cm)was significantly greater (t 5 8.44, df 5 33, P ,
0.0001; Table 3) than paired randomly-selectedtrees (48.34 cm). Mean height of nest-treeemergence (5.29 m) above the surroundingcanopy was significantly greater (t 5 3.44, df 5
33, P 5 0.0016; Table 3) than the mean height ofrandomly-selected tree emergence (0.70 m). Themean (6 SE) height of Mississippi Kite nests was26.89 6 0.82 m, while the mean surroundingcanopy height was 27.68 6 0.74 m, indicatingkites tended to build nests almost level with theheight of the surrounding canopy.
The mean (6 SE) distance (61.06 6 15.65 m)to the forest edge from Mississippi Kite nests wassignificantly less (t 5 4.16, df 5 33, P 5 0.0002;Table 3) than the distance (126.19 6 16.18 m) tothe forest edge from randomly-selected sites.Other measured variables did not differ betweennest and random sites (Table 3).
Twenty of 35 nest sites (57%) were in crotchesof secondary branches. The remaining 15 nestswere on primary branches of nest trees. Nestswere randomly placed in any direction throughoutthe nest trees. Mississippi Kite nests were found insix species of trees with Overcup oak (Quercuslyrata; n 5 11) used most commonly. Sweetgum(Liquidamber styraciflua; n 5 7), willow oak (Q.phellos) (n 5 7), Nuttall oak (Q. nuttallii; n 5 3),American sycamore (Platanus occidentais; n 5
2), and white oak (Q. alba; n 5 1) were used lessfrequently as nesting sites.
Mississippi Kites used numerous species oftrees for perching. Black willow (Salix nigra; n 5
9) and water hickory (Carya aquatica; n 5 8)were the two most common species used forprimary perches. Overcup oak (n 5 5) and Nuttalloak (n 5 4) were also commonly used for primaryperches. Sweetgum (n 5 3), slippery elm (Ulmusrubra; n 5 3), willow oak (n 5 2), and water oak(Q. nigra; n 5 2) were also used as primary perchtrees. Nineteen (52.8%) of 36 primary perch trees(Table 4) were live with a dead limb used as theperch, six were live trees with sparsely-leavedlimbs, six were live trees with live limbs used asperches, and five were dead trees.
Secondary perch trees included water hickory(n 5 3), overcup oak (n 5 2), water oak (n 5 1),and sugarberry (Celtis laevigata; n 5 1). Onedead tree, three live trees with sparsely-leaved
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:13 710 Cust # 09-121
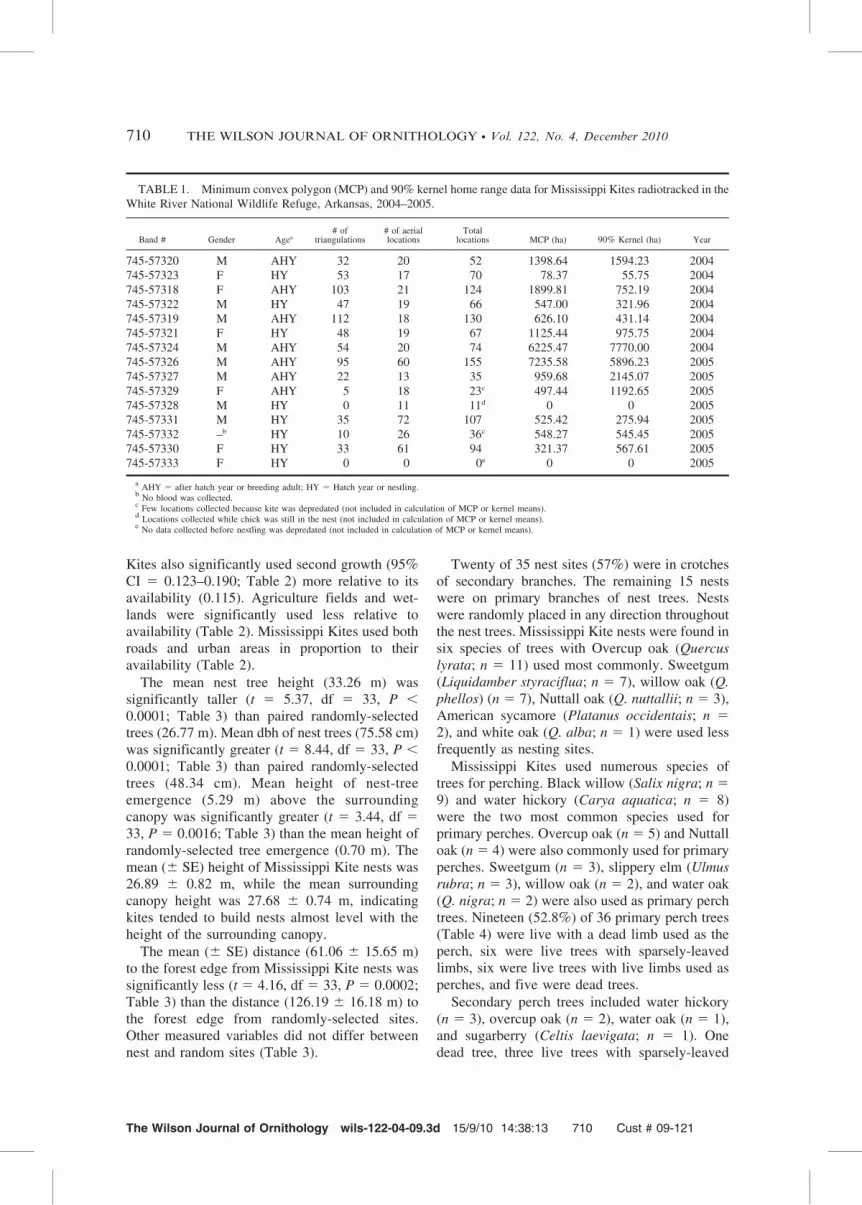
TABLE 1. Minimum convex polygon (MCP) and 90% kernel home range data for Mississippi Kites radiotracked in the
White River National Wildlife Refuge, Arkansas, 2004–2005.
Band # Gender Agea# of
triangulations# of aeriallocations
Totallocations MCP (ha) 90% Kernel (ha) Year
745-57320 M AHY 32 20 52 1398.64 1594.23 2004
745-57323 F HY 53 17 70 78.37 55.75 2004
745-57318 F AHY 103 21 124 1899.81 752.19 2004
745-57322 M HY 47 19 66 547.00 321.96 2004
745-57319 M AHY 112 18 130 626.10 431.14 2004
745-57321 F HY 48 19 67 1125.44 975.75 2004
745-57324 M AHY 54 20 74 6225.47 7770.00 2004
745-57326 M AHY 95 60 155 7235.58 5896.23 2005
745-57327 M AHY 22 13 35 959.68 2145.07 2005
745-57329 F AHY 5 18 23c 497.44 1192.65 2005
745-57328 M HY 0 11 11d 0 0 2005
745-57331 M HY 35 72 107 525.42 275.94 2005
745-57332 –b HY 10 26 36c 548.27 545.45 2005
745-57330 F HY 33 61 94 321.37 567.61 2005
745-57333 F HY 0 0 0e 0 0 2005
aAHY 5 after hatch year or breeding adult; HY 5 Hatch year or nestling.
bNo blood was collected.
cFew locations collected because kite was depredated (not included in calculation of MCP or kernel means).
dLocations collected while chick was still in the nest (not included in calculation of MCP or kernel means).
eNo data collected before nestling was depredated (not included in calculation of MCP or kernel means).
710 THE WILSON JOURNAL OF ORNITHOLOGY N Vol. 122, No. 4, December 2010
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:15 711 Cust # 09-121
FIG. 2. Minimum Convex Polygon (MCP) and 90% kernel home range of a radio-tracked Mississippi Kite (Band #
745-57331) in the White River National Wildlife Refuge, Arkansas, 2004–2005.
Bader and Bednarz N HOME RANGE AND HABITAT OF MISSISSIPPI KITES 711
limbs, two live trees with a dead limb, and onelive tree with live limbs were all used assecondary perch trees.
There were no significant differences in dbh,tree height, distance to edge, or distance to nestbetween primary and secondary perch trees(Table 4). Primary and secondary perch treeswere significantly closer (t 5 5.6, df 5 37, P 5
,0.001) to an edge and had a significantly larger(t 5 7.5, df 5 34, P , 0.0001) dbh than treesselected at random (Tables 3, 4). There was nosignificant difference in height of primary andsecondary trees and trees selected at random.
DISCUSSION
The mean 90% kernel home range size of maleMississippi Kites (3,567 ha, n 5 5) was largerthan the one female monitored (752 ha). Wesuggest this difference may be real because malesare more likely to venture long distances from thenest during their daily foraging activities (Dunstanet al. 1978, Barber 1995). Males range widely,probably to gain access to habitat types or food
resources that are not available near the nest site.
Access to a larger foraging area and possibly to
areas with more abundant prey makes the male’s
role as a food provider to the nestlings vital to
success of the nest. Female kites may have smaller
home ranges because of their role in the primary
defense of the nest; they also tend to the nestlingsmore than males, similar to other female raptors
(Newton 1978, Collopy 1984, Bader 2007).
Barber (1995) estimated the mean male MCP
Mississippi Kite home range size in Missouri as
1,357 ha (n 5 6), notably smaller than the mean
male MCP home ranges that we estimated
(3,289 ha). The mean female kite home range in
Missouri was 865 ha (n 5 4; Barber 1995), also
considerably smaller than the MCP home range of
the one female that we radiotracked (1,900 ha).
We suggest these difference may be due to either
differences in landscapes of the respective study
areas (the Missouri study was conducted in a more
fragmented, forest-agriculture landscape) and the
techniques used (no aerial telemetry was used in
the Missouri study), or both factors.
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:21 712 Cust # 09-121
TABLE 3. Habitat characteristics at Mississippi Kite nest sites (n 5 39) and sites selected at random (n 5 39) in the
White River National Wildlife Refuge, Arkansas, 2004–2005.
* Significantly different from sites selected at random, P # 0.05.a
P 5 probability that means are different based on a paired t-test.b
Mean tree height 5 mean height of trees in a 0.04-ha plot surrounding the nest or random tree.c
Tree emergence 5 the difference between mean height of nest trees above surrounding trees in 0.04 ha plot or mean height of random trees above surroundingtrees in 0.04-ha plot.
TABLE 2. The proportion of cover types within home ranges available and used by radio-tracked adult (n 5 6) and
juvenile (n 5 6) Mississippi Kites in the White River National Wildlife Refuge, Arkansas, 2004–2005.
Second growth 2417.41 136 0.115 0.155* 0.123–0.190
Road 64.83 2 0.003 0.002 0.000–0.011
Wetland 821.65 13 0.039 0.015* 0.006–0.029
Urban 38.59 6 0.002 0.007 0.001–0.018
Water 1488.13 88 0.071 0.100* 0.074–0.130
* Significantly different from amount available.
712 THE WILSON JOURNAL OF ORNITHOLOGY N Vol. 122, No. 4, December 2010
Our data documented that Mississippi Kitessignificantly used open waterways more thanavailable to kites within their home ranges(Table 2). Open water likely provided habitat forprey items such as dragonflies (Odonata) andfrogs (Anura). Barber (1995) did not report thiscover type to be more abundant within kite homeranges compared to reference areas, but it was alarge part of most home ranges that he sampled.
Agricultural fields were used less within thekite home ranges in Arkansas than available(Table 2), but were significantly more abundantin kite home ranges in Missouri (Barber 1995).Soybeans, milo, corn, cotton, and fallow fieldswere included in the agriculture cover type in theMissouri study. Rice, soybeans, and cotton werethe primary crops farmed in the areas surroundingthe WRNWR.
Wetland areas were significantly underused(Table 2), but we suggest wetlands indirectlymay be an important area for foraging kites.Wetlands likely produce prey, but this prey is notrestricted to these areas and probably moves intoother cover types where kites may forage on them.Wet areas are used as foraging habitats by otherraptors such as the Red-shouldered Hawk (Bed-narz and Dinsmore 1981, Bloom et al. 1993,Howell and Chapman 1997).
Mature forests were significantly used morewithin the home ranges of Mississippi Kites inMissouri (Barber 1995) and in Arkansas (Table 2).In Tennessee, Kalla (1979) reported wooded areasin the floodplain provided preferred nesting sites forkites and key habitat for prey species. Matureforests provide habitat for dragonflies and cicadas(Homoptera), the two most common prey itemsdelivered to nests in the WRNWR (Bader 2007).Red-shouldered Hawks were demonstrated to useforested habitats in California (Bloom et al. 1993)and bottomland hardwood forests in Georgia(Howell and Chapman 1997) more than available.Large contiguous tracts of mature forests seem to be
an important cover type for kites providing bothnest sites and prey.
Second growth was used significantly morewithin home ranges of kites in Arkansas (Table 2),but not in Missouri (Barber 1995). This cover typeis common in numerous landscapes, and canprovide foraging habitat and marginal nestinghabitat for kites. Timber stands classified as secondgrowth can provide nest sites if super-emergenttrees are not removed during timber harvestoperations or are created during timber harvest.Fallow fields, which consist of early successionalgrowth, can provide habitat for aerial insects andother prey items. The second growth cover type wasnot common in kite home range in Missouri andonly accounted for 3.8% of home ranges and 1.2%
of the sampled surrounding areas.
Kites used urban areas and roads in proportionto availability within their home range. Bothurban areas and roads were near other cover typesused by kites. Kites may have used thermalsproduced in these areas and probably did notactively select these sites for foraging.
Mississippi Kites selected trees taller than themean overstory canopy height in their nesting areas(Table 3). Use of super-emergent trees for nestingallows this long-winged raptor efficient access tothe nest (Barber et al. 1998). Kites in WRNWRtended to build nests almost level with the top ofthe surrounding canopy. Barber (1995) and Whit-mar (1987) reported nests were higher than the topsof the surrounding trees. Placing nests at or slightlyabove the tops of the surrounding trees mayprovide a view of the surrounding area andpotential predators (Whitmar 1987, Barber 1995).Placing nests slightly below the surroundingcanopy height would give the nest more protectionfrom the wind and make the nest less conspicuousto predators. Mississippi Kite nest trees have alarger dbh than randomly-selected trees (Table 3).Barber (1995) also reported the dbh of nest trees tobe significantly larger than random trees. The large
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:22 713 Cust # 09-121
TABLE 4. Characteristics of primary (n 5 36) and secondary (n 5 7) perch trees of Mississippi Kites in the White
River National Wildlife Refuge, Arkansas, 2004–2005.
dbh of nests trees may be correlated with age andsize of the super-emergent trees typically used fornesting sites.
Mississippi Kites selected nest sites closer tothe forest edge than random sites and all forestedges comprised of lakes, bayous, or the WhiteRiver. This pattern was also documented duringresearch previously conducted on the refuge (St.Pierre 2006). These bodies of water provide openareas for kites to forage for aerial insects. Barber(1995) reported kite nests were located closer tofield edges more frequently than water edges.Field edges were uncommon within the interior ofWRNWR, where waterways comprised the ma-jority of the available edges. Whitmar (1987)reported kite nests in the Mississippi River Valleyin Missouri had a mean distance to an edge of49.2 m and a mean distance to water of 96.3 m.Edge habitats, particularly natural edges, seem tobe an important attribute of Mississippi Kitenesting habitat.
Kite nests were more commonly built onsecondary than on primary branches. Barber(1995) also documented this pattern for kite nestsin Missouri and suggested this may be an anti-predator defense. Our data and those reported byWhitmar (1987) indicated kite nests were seem-ingly randomly placed throughout the nest tree.
CONSERVATION IMPLICATIONS
Mature tracts of forests should be conservedbecause this habitat type provides nesting sitesand a foraging habitat for Mississippi Kites.Large, super-emergent trees should be conservedfor kite nesting habitat. Management that pro-motes development of additional large, super-emergent trees will likely be beneficial to kites.Timber harvests consistent with kite conservationshould be conducted so uneven-height canopyforests are created. We recommend that second-growth forest should be allowed to develop longerbetween harvests (.75 years) to provide foragingand nesting habitat for kites. Managers shouldavoid even-aged regeneration of clear-cuts andfallow fields. Natural waterways, including wet-lands, should be conserved for production of preyand as foraging areas.
ACKNOWLEDGMENTS
This project was funded primarily by the U.S. Fish and
Wildlife Service (USFWS) and Arkansas Game and Fish
Commission (AGFC) through a State Wildlife Grant.
Substantial additional funds were provided by Arkansas
State University. We thank K. L. Rowe (AGFC) for support
and assistance on this project. We thank the USFWS,
especially R. E. Hines and staff at the White River National
Wildlife Refuge, for help on the logistics for this project.
We are especially grateful to our research technician, W. D.
Edwards for hard work and dedication to this project. We
greatly appreciate the assistance in the field and in the
laboratory provided by J. D. Brown, T. J. Benson, D. J.
Baxter, T. R. Edwards, E. J. Bader, and A. M. St. Pierre.
LITERATURE CITED
ALLAN, P. F. AND P. R. SIME. 1943. Distribution and
abundance of the Mississippi Kite in the Texas
panhandle. Condor 45:110–112.
BADER, T. J. 2007. Reproductive success, causes of nesting
failures, and habitat use of Swallow-tailed and
Mississippi kites in the White River National Wildlife
Refuge, Arkansas. Thesis. Arkansas State University,
Jonesboro, USA.
BADER, T. J. AND J. C. BEDNARZ. 2009. Reproductive
success and causes of nesting failures for Mississippi
Kites: a sink population in eastern Arkansas? Wetlands
29:598–606.
BAILEY, B. J. R. 1980. Large sample simultaneous
confidence intervals for the multinomial probabilities
based on transformation of the cell frequencies.
Technometrics 22:583–589.
BARBER, J. D. 1995. Mississippi Kite habitat use and
reproductive biology in the Mississippi flood plains of
southeast Missouri. Thesis. University of Missouri,
Columbia, USA.
BARBER, J. D., E. P. WIGGERS, AND R. B. RENKEN. 1998.
Nest-site characterization and reproductive success of
Mississippi Kites in the Mississippi River floodplains.
Journal of Wildlife Management 62:1373–1378.
BEDNARZ, J. C. AND J. J. DINSMORE. 1981. Status, habitat
use, and management of Red-shouldered Hawks in
Iowa. Journal of Wildlife Management 45:236–241.
BLOOM, P. H., M. D. MCCRARY, AND M. J. GIBSON. 1993.
Red-shouldered Hawk home-range and habitat use in
southern California. Journal of Wildlife Management
57:258–265.
CHERRY, S. 1996. A comparison of confidence interval
methods for habitat use-availability studies. Journal of
Wildlife Management 60:653–658.
COLLOPY, M. W. 1984. Parental care and feeding ecology
of Golden Eagle nestlings. Auk 101:753–760.
DEVAULT, T. L., W. L. STEPHENS, B. D. REINHART, O. E.
RHODES JR., AND I. L. BRISBIN JR. 2003. Aerial
telemetry accuracy in a forested landscape. Journal of
Raptor Research 37:147–151.
DONOHUE, K. C. AND A. M. DUFTY JR. 2006. Sex
determination of Red-tailed Hawks (Buteo jamaicensis
calurus) using DNA analysis and morphometrics.
Journal of Field Ornithology 77:74–79
DUNSTAN, T. C., J. H. HARPER, AND K. B. PHIPPS. 1978.
Habitat use and hunting strategies of Prairie Falcons,
Red-tailed Hawks, and Golden Eagles. Final Report
submitted to USDI, Bureau of Land Management,
Denver, Colorado, USA.
The Wilson Journal of Ornithology wils-122-04-09.3d 15/9/10 14:38:23 714 Cust # 09-121
714 THE WILSON JOURNAL OF ORNITHOLOGY N Vol. 122, No. 4, December 2010